JAK MÓZG WIDZI BARWY?
Szczypta filozofii na początek
Próbując odpowiedzieć na tytułowe pytanie: „Jak mózg widzi barwy?” nie można uchylić się od odpowiedzi na znacznie bardziej podstawowe pytanie dotyczące barwności tego, co miałby widzieć mózg, czyli świata. Innymi słowy, rzecz w tym, czy otaczająca nas rzeczywistość jest kolorowa. Codzienne doświadczenie nieustannie utwierdza nas w tym przekonaniu. Wielobarwność świata nie jest jednak tak oczywista, jak mogłoby to wyglądać na pierwszy rzut oka. Przede wszystkim, jeśli chcemy mówić coś o otaczającym nas świecie musimy najpierw zgodzić się co do tego, gdzie świat się kończy, a gdzie się zaczyna ten, który go poznaje. Kluczowe w tej kwestii jest rozstrzygnięcie, w którym miejscu leży granica między światem-który-mnie-otacza a ja-widzącym. Ostatecznie to przecież ja-widzące doświadcza barwności świata i coś o nim twierdzi.
Nie jest moją intencją uprawianie tutaj jakieś naiwnej filozofii, ale raczej zasygnalizowanie problemu, który ma istotne znaczenie dla rozumienia czym jest percepcja. Otóż, najbardziej oczywista odpowiedź, jaka przychodzi na myśl w sprawie granicy między ja-widzącym a otaczającym-mnie-światem opiera się na spostrzeżeniu, że tym, co oddziela mnie od świata jest po prostu moja skóra. Wszystko to, co znajduje się poza nią, nie jest mną, a zatem należy do świata, który poznaję. Paradoks polega na tym, że przyjmując takie założenie, jako oczywiste musimy odrzucić inne, równie oczywiste.
Otóż, jeżeli granicą oddzielającą mnie od świata jest moja skóra to świat jest bezbarwny. Trawa nie jest zielona, a niebo nie jest niebieskie. To czysta iluzja, jaką fundują mi oczy i mózg, które się znajdują przecież w skórzanym worku mojego ciała. Co więcej, z tego punktu widzenia świat nie tylko nie jest bezbarwny, ale gorzej – nawet nie jest szarobury! Po prostu kategorie jasności, podobnie, jak kategorie barwności nie są własnościami światła tylko tego, kto jest na nie wrażliwy.
Wystarczy się chwilę zastanowić nad tym, co zostało już zasygnalizowane na stronach niniejszej książki, żeby zrozumieć tę oczywistość. Tym co pobudza fotoreceptory w siatkówkach oczu jest światło widzialne, a właściwie fala elektromagnetyczna o długości w zakresie ok. 400–700 nanometrów i zmiennej amplitudzie . I tylko tyle wiemy o tej fali. Chociaż barwę kojarzymy z taką czy z inną długością światła, tym niemniej ono samo nie jest ani jasne ani ciemne, ani tym bardziej barwne. To po prostu niektóre fotoreceptory reagują intensywniej na określoną długość i amplitudę tej fali, a mózg interpretuje ich reakcję, tworząc wrażenie barwy o określonej intensywności i jasności.
Oczywiście nie chciałbym wyjść na nudziarza, ale jednak przypomnę raz jeszcze, że jak dotąd nie mamy zielonego pojęcia, jak na podstawie reakcji milionów komórek nerwowych zaangażowanych w przetwarzanie danych zarejestrowanych przez fotoreceptory dochodzi do wrażenia widzenia barwy. To zagadnienie zostawmy do wyjaśnienia profesjonalnym filozofom. W każdym razie, nauki empiryczne nie stawiają na razie żadnych weryfikowalnych hipotez w tej sprawie.
Jeśli jednak nie podoba nam się idea bezbarwności świata, to pozostaje zastanowić się nad tym czy nie dałoby się jakoś przesunąć granicy między ja-widzącym a otaczającym-mnie-światem, żeby rzeczywistości znajdującej się poza mną przydać nieco blasku. Dla naszych potrzeb raczej trudno byłoby uzasadnić poszerzanie naszego ja-widzącego poza skórę, skoro raczej nie mamy powodów, żeby wątpić, iż to oczy i mózg tworzą barwy. Należałoby więc raczej przesuwać granicę ja-widzącego do wnętrza ciała, aby te struktury mózgu, które są odpowiedzialne za widzenie barwne znalazły się po stronie świata. Wtedy już nikt nie mógłby zakwestionować oczywistego przecież doświadczenia jego barwności. Niestety takie postawienie sprawy tylko piętrzy kolejne problemy. Na dwa z nich zwrócę uwagę.
Po pierwsze, wszystkie stany umysłowe, w tym również doświadczenie samego siebie, czyli także ja-widzącego, są w mniejszym lub większym stopniu pochodną sprawnie działającego całego mózgu. Jak zatem oddzielić od siebie te jego części, które są odpowiedzialne za widzenie barwne od tych, które leżą u podłoża naszego odczuwania ja? I po drugie, wprowadzenie do naszego rozumowania jakiegoś niematerialnego lub poza-neuronalnego pierwiastka, który byłby odpowiedzialny za istnienie ja (w każdej z jego ról), w opozycji do stanów umysłu będących skutkiem jego aktywności poznawczej, np. widzenia barwności świata, wyrzuciłoby nas poza granice nauki empirycznej w znaczeniu, jakie współcześnie przynależy temu pojęciu.
Chyba nie ma zatem sensu dalsze rozwijanie tych wątków, ponieważ znacznie wykroczyłyby one poza ramy tej książki. Dość powiedzieć, że koncepcja, zgodnie z którą świat jest bezbarwny i bez-jasny, a granica naszego ja-widzącego znajduje się na rogówce oka, wydaje się wyjściem najrozsądniejszym, przynajmniej na obecnym etapie wiedzy o widzeniu.
Wypełnianie konturów czy konturowanie barwnych plam?
Na podstawie tego, co do tej pory napisałem w odniesieniu do spostrzegania konturów widzianych rzeczy można sformułować ogólną zasadę, że obiekty w scenie wizualnej są tym trafniej identyfikowane, im wyraźniej jawią się nam ich kontury wyznaczone przez kontrastujące ze sobą powierzchnie o różnej jasności, im bardziej te kontury są kompletne i im mniej są „zaszumione”, np. światłocieniami.
Mike G. Harris i Glyn W. Humphreys (2002) zauważają, że poproszeni o narysowanie jakiegokolwiek przedmiotu najczęściej zaczynamy od wykreślenia jego konturów, a nie, np. od zacienienia lub zabarwienia kilku leżących obok siebie powierzchni. Pomijając już fakt, że w znacznym stopniu odpowiada za to mechanizm hamowania obocznego ukształtowany w toku ewolucji, tym niemniej od dziecka jesteśmy trenowani raczej w wypełnianiu konturowych obrazków różnymi kolorami (jak, np. na ryc. 107), niż obrysowywaniu konturami barwnych powierzchni. I czynimy tak ignorując fakt, że zdecydowana większość przedmiotów nie ma na sobie żadnych linii konturowych. Że to, co widzimy to są właśnie wyłącznie różnobarwne, a w słabszych warunkach oświetleniowych, tylko różno-jasne powierzchnie, które wyznaczają granice przedmiotów oddzielających je od tła.

Jaką rolę odgrywają zatem barwy w spostrzeganiu otaczającego nas świata? Czy potrzebujemy ich do identyfikacji przedmiotów? A może widzenie barw jest tylko ekstra-dodatkiem, w jaki wyposażyła nas ewolucja?
Z jednej strony, stosunek ilości czopków, które są odpowiedzialne za widzenie barwne, do ilości pręcików, które reagują na jasność światła w słabszych warunkach oświetleniowych, wynosi ok. 1:20. Oznacza to, że rzeczywiście można mieć wątpliwość czy widzenie barwne jest nam do czegokolwiek potrzebne. Z drugiej jednak strony, w okolicach dołka centralnego, czyli miejsca szczególnego ze względu na ostrość widzenia, znajduje się nieporównywalnie więcej czopków niż pręcików. Wobec tego można słusznie przypuszczać, że widzenie zapośredniczone wrażliwymi na barwy czopkami odgrywa jednak jakąś szczególną rolę w oglądaniu świata.
Wszystko wskazuje na to, że bez przyjrzenia się mechanizmom odpowiedzialnym za widzenie barwne nie rozwiążemy tych dylematów. Warto rozpocząć tę opowieść od przypomnienia kilku faktów z historii.
Trójchromatyczna teoria Younga-Helmholtza
Trójchromatyczna teoria widzenia barw (the trichromatic color theory) powstała w XVIII wieku, a jej prekursorami byli Brytyjczycy, Sir Isaak Newton, George Palmer i Sir Thomas Young. Pierwszy z nich położył kamień węgielny zarówno pod współczesną optykę, jak i teorię widzenia barwnego. Wyniki jego eksperymentów nad rozszczepieniem światła białego na spektrum barw (colour spectrum) zanegowały wielowiekową tradycję, zgodnie z którą to nie światłu, ale przedmiotom przypisywano barwność.
Dotąd uważano, że barwa jest własnością rzeczy, a nie wzroku. Sądzono, że światło wydobywa barwy z przedmiotów i dzięki temu są one spostrzegane przez obserwatora. Wszystkie koty są szare po zmroku właśnie dlatego, że jest zbyt mało światła, aby ujawniło się ich faktyczne umaszczenie. Wystarczy tylko zapalić lampę i natychmiast widać, że są, np. rude. Newton zakwestionował tę ideę. Zwrócił uwagę na to, że ten sam obiekt, w zależności od intensywności i jakości, czyli odcienia padającego nań światła może wywoływać zupełnie inne wrażenia barwne. Na tej podstawie wyciągnął wniosek, że widzenie barwne jest w znacznie większym stopniu pochodną działania systemu wzrokowego, niż własności oglądanych przedmiotów.
Zafascynowany odkryciami Newtona, George Palmer w opublikowanej w 1777 roku książce zasugerował, że włókna nerwowe, z których zbudowana jest siatkówka oka zawierają trzy rodzaje cząsteczek barwnikowych (coloring particles). Każda z tych cząsteczek pochłania promienie światła o długości odpowiadającej jednej z barw: czerwonej, żółtej lub niebieskiej. Według Palmera widzenie różnych barw jest wynikiem mieszania się dwóch lub trzech cząsteczek barwnikowych w oku w takiej proporcji, w jakiej pochłaniają one światło o tej długości, na jaką są wrażliwe. Palmer był chemikiem i dlatego mechanizm widzenia barw tłumaczył przez analogię do procesu mieszania farb, czyli tzw. syntezy subtraktywnej (subtractive color; ryc. 108).
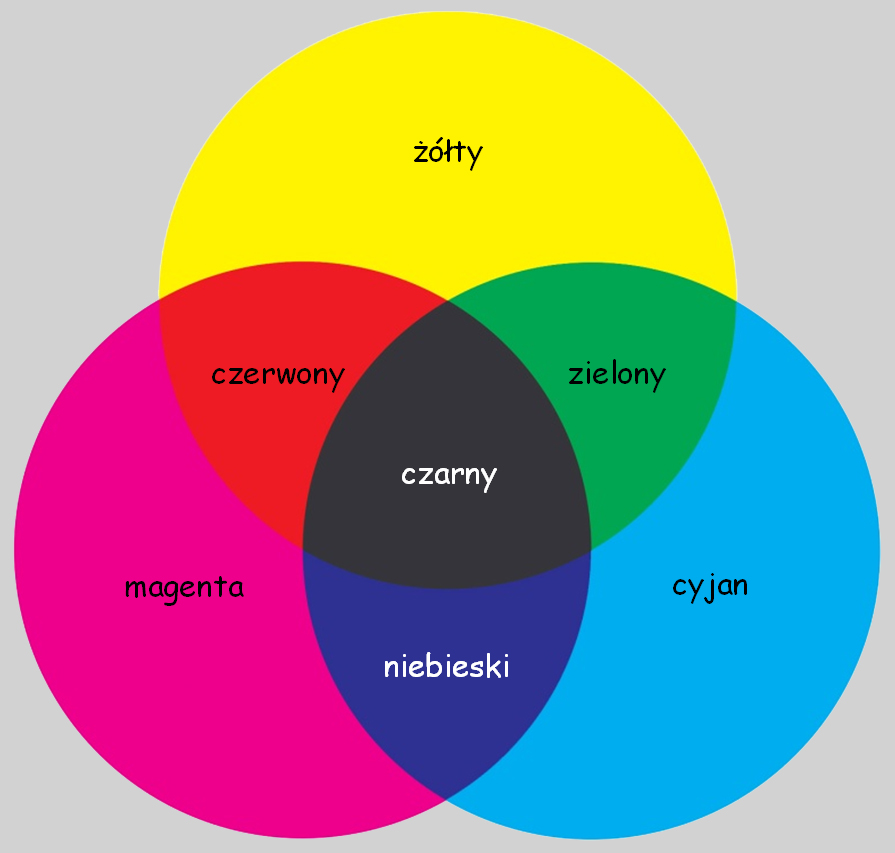
Zdaniem Palmera, wrażenie barwy zielonej powstaje w rezultacie zmieszania się dwóch cząstek barwnikowych, które pochłonęły promienie światła o długości odpowiadającej barwie żółtej i cyjanu, czyli odcienia barwy niebieskiej. Czerwień jest pochodną mieszaniny barwników pochłaniających światło o długości odpowiadającej barwie żółtej i magenty, a niebieski – pochodną magenty i cyjanu. Światło pochłaniane przez wszystkie trzy rodzaje cząstek daje wrażenie czerni, zaś biel jest konsekwencją braku pobudzenia któregokolwiek z siatkówkowych barwników (Lowengard, 2006).
Georges Palmer zbudował swoją koncepcję na dwóch filarach: newtonowskiej optyce oraz teorii mieszania kolorów, opracowanych przez francuskich chemików, Charlesa Dufay i Jean Hellota. Jego koncepcja przetrwała co prawda przez następnych 30 lat, ale budziła wiele kontrowersji. Przede wszystkim Palmer nie umiał wyjaśnić dlaczego postulowane przez niego cząstki barwnikowe miałyby reagować tylko na trzy wskazane przez niego długości fali świetlnej. Czyżby pozostałe długości fali światła widzialnego nie odgrywały żadnej roli w widzeniu barwnym? A po drugie, wraz z rozwojem fizjologii coraz większe wątpliwości budziła jego koncepcja włókien nerwowych, w których – zdaniem Palmera – niczym w maleńkich tygielkach mieszałyby się trzy cząstki barwnikowe.
W 1807 roku, Sir Thomas Young, podczas wykładów z filozofii naturalnej, przedstawił koncepcję, zgodnie z którą siatkówka oka nie jest zbudowana z włókien nerwowych wypełnionych trzema cząsteczkami barwnikowymi, ale z trzech rodzajów komórek receptorowych, które z różną intensywnością reagują na promienie świetlne obejmujące cały zakres światła widzialnego. Young sugerował także, aby w rozważaniach nad widzeniem barwnym odstąpić od idei mieszania się barw w oku, tak jak przedstawił to Palmer, odwołując się do analogii z mieszaniem farb. W zamian Young, postulował koncepcję sumowania się siły reakcji receptorów na pobudzenie falą świetlną o różnej długości. Chociaż generalnie zgadzał się z przyjętą przez Palmera ideą, zgodnie z którą wszystkie barwy są pochodną mieszania trzech składników, tym niemniej jako podstawę do ich wyznaczenia przyjął spectrum barw uzyskane w wyniku rozszczepienia światła białego. Trzy punkty maksymalnej wrażliwości receptorów związał on z barwą niebieską, zieloną i czerwoną (ryc. 109). Wybór, jakiego dokonał był oczywiście arbitralny, ale nie pozbawiony logiki. Po podziale całego spectrum światła widzialnego na trzy części, wyróżnione przez niego barwy zdecydowanie dominują w ramach każdej kategorii.

Warto w tym miejscu dodać, że chociaż wyniki późniejszych badań fizjologicznych nie w pełni potwierdziły intuicję Younga, dotyczącą wrażliwości fotoreceptorów na barwę czerwoną, zieloną i niebieską, tym niemniej ich podział na czopki typu R (red), G (green) i B (blue) utrzymuje się do dzisiaj.
Modelowy opis zależności zachodzących między długością fali świetlnej a wrażliwością postulowanych przez Younga trzech rodzajów fotoreceptorów przedstawił w 1851 roku niemiecki fizjolog i fizyk, Hermann von Helmholtz. Podobnie, jak Young, twierdził on, że widzenie różnych barw wiąże się ze zróżnicowaną wrażliwością fotoreceptorów na wszystkie długości fali światła widzialnego. Uważał również, że dla każdego typu fotoreceptora istnieje taka długość fali świetlnej, na którą jest on najbardziej wrażliwy (zob. ryc. 110).
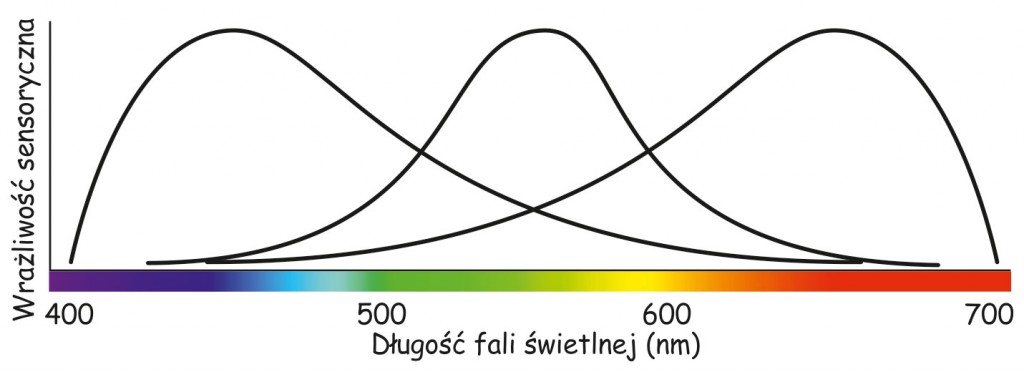
Z wykresu na ryc. 110 wynika, że widzenie barwy, np. pomarańczowej jest związane z dużą aktywnością receptorów wrażliwych na długie fale światła widzialnego, znacznie mniejszą, na średnie długości fali świetlnej i śladową, na najkrótsze fale świetlne. Z kolei widzenie, np. barwy zielonej jest związane z maksymalną aktywnością receptorów wrażliwych na średnią długość fali świetlnej i nieznaczną aktywność dwóch pozostałych typów fotoreceptorów. Zjawisko mieszania barw w wyniku sumowania się fal świetlnych o różnej długości nazywa się syntezą addytywną (additive color) i podobnie, jak koncepcja subtraktywnego mieszania barwników, pozwala przewidzieć, jaka barwa powstanie z połączenia dwóch lub trzech składników światła o różnej barwie (ryc. 111).
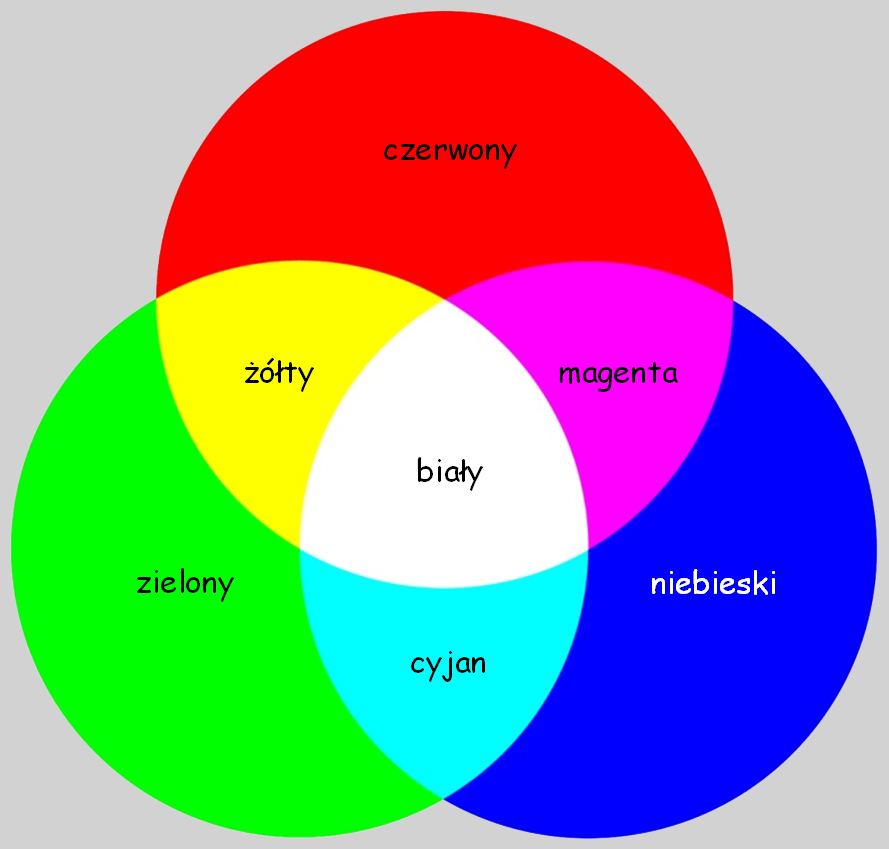
Tym razem wrażenie barwy żółtej powstaje w rezultacie zmieszania się dwóch strumieni światła o długościach odpowiadających barwie czerwonej i zielonej. Magenta jest barwą wynikową z dwóch strumieni światła, których długość odpowiada barwie czerwonej i niebieskiej, natomiast wrażenie cyjanu powstaje na skutek oświetlenia płaszczyzny światłem o długości odpowiadającej barwie niebieskiej i zielonej.
Opracowana przez Younga i Helmholtza koncepcja trzech typów receptorów wrażliwych na falę świetlną o różnej długości doczekała się empirycznego potwierdzenia dopiero w 1956 roku przez Gunnara Svaetichina. Wykazał on, że w siatkówce oka ryby rzeczywiście znajdują się trzy rodzaje fotoreceptorów (tzw. czopków), które są szczególnie wrażliwe na trzy różne długości fali świetlnej, mniej więcej odpowiadające postulowanym przez Younga i Helmholtza barwom: niebieskiej, zielonej i czerwonej. Kilka lat później podobne wyniki uzyskali biofizycy z Johns Hopkins University (Marks, Dobelle i MacNichol, 1964) podczas badań nad fotoreceptorami znajdującymi się w siatkówce oka u małp i w tym samym roku, biolodzy z Harvard University (Brown i Wald, 1964) – w odniesieniu do fotoreceptorów znajdujących się w siatkówce oka człowieka.
Czopki R, G i B czy L, M i S?
Faktyczne zakresy intensywności reakcji trzech hipotetycznych rodzajów czopków na falę elektromagnetyczną w zakresie światła widzialnego ilustruje ryc. 112. Na wykresie zaznaczono także zakresy reaktywności pręcików, czyli fotoreceptorów aktywnych w warunkach obniżonej jasności. Nawiasem mówiąc ani Young, ani Helmholtz nie przypuszczali, że w oku mogą znajdować się dwa rodzaje fotoreceptorów, z których jeden rodzaj jest wrażliwy na intensywne światło dzienne, a drugi – aktywny w warunkach nocnych. Zasadniczo ich koncepcja dotyczyła tylko pierwszego rodzaju receptorów, czyli czopków. Maksymalna wrażliwość pręcików przypada na falę światła widzialnego o długości ok. 500 nm i stosunkowo niskiej intensywności (prawie trzykrotnie niższej niż intensywność światła, na które reagują czopki). Z kolei w grupie czopków występują wspomniane już trzy rodzaje fotoreceptorów, wrażliwych na różne długości fali świetlnej, które są interpretowane przez mózg, jako barwy: czerwona, zielona i niebieska.
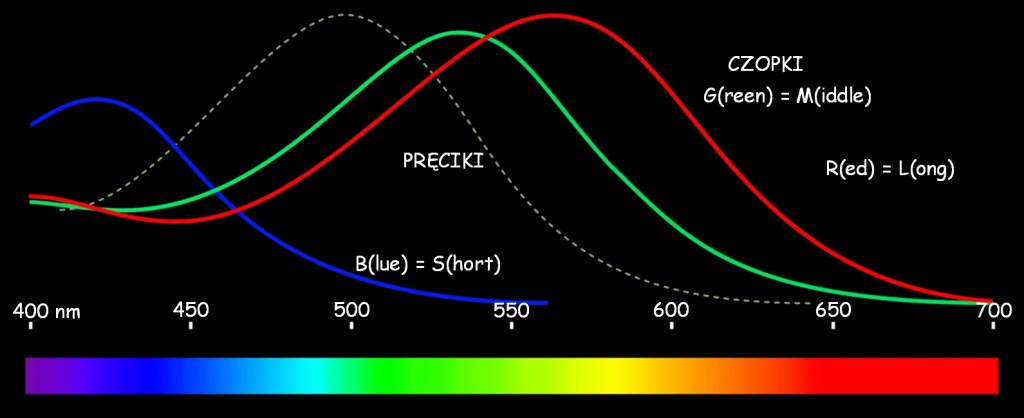
Czopki typu R najintensywniej reagują na falę światła widzialnego o długości ok. 560 nm. Ponieważ spośród wszystkich fotoreceptorów są wrażliwe na najdłuższą falę elektromagnetyczną w zakresie światła widzialnego, dlatego zwane są również czopkami typu L (long). Niezupełnie ta długość fali odpowiada barwie czerwonej, ale wrażenie czerwieni rzeczywiście powstaje w wyniku działania tego fotoreceptora tylko w nieco bardziej skomplikowany sposób, niż opisuje to teoria Younga-Helmholtza. Będzie o tym jeszcze mowa. Na falę elektromagnetyczną o długości ok. 530 nm, najintensywniej reagują czopki typu G. Ponieważ ich maksymalna wrażliwość przypada na falę światła widzialnego o długości pośredniej między długą i krótką, dlatego oznacza się je również literą M (middle). Z kolei na światło niebieskie o długości ok. 420 nm najintensywniej reagują czopki typu B lub, inaczej, typu S (short), ponieważ ich maksymalna wrażliwość jest związana z najkrótszymi długościami fali światła widzialnego (Matthews, 2000).
Czopki typu R, G i B nie są równolicznie reprezentowane w siatkówce. Najmniej jest czopków reagujących na najkrótszą falę elektromagnetyczną w zakresie światła widzialnego, czyli na barwę niebieską. Stanowią one tylko 5–6% z ok. 5 milionów czopków. Ta proporcja jest względnie stała u wszystkich ludzi. Co więcej, czopków typu B nie ma w dołku centralnym, a na pozostałym obszarze siatkówki występują bardzo nieregularnie (Hofer, Carroll i Williams, 2009). Nie bardzo wiadomo dlaczego tak jest, ale bynajmniej nie przeszkadza to w normalnym widzeniu barwy niebieskiej.
Z kolei czopków typu R, reagujących najintensywniej na długie fale światła widzialnego, na ogół jest znacznie więcej niż czopków typu G, które reagują najintensywniej na średnią długość fali światła widzialnego, chociaż okazuje się, że istnieją znaczne różnice w ich proporcji u różnych ludzi (Hofer, Carroll, Neitz i Williams, 2005). Zachowując normalną zdolność do różnicowania barwy czerwonej i zielonej, u niektórych osób proporcja liczby czopków typu R do G może wynosić od 1,9:1 do 16,5:1, ale są i tacy, u których jest ona odwrócona, np. 0,37:1 (ryc. 113). Tak czy inaczej, w samym dołku centralnym występują tylko czopki typu R i G.

Zakres widzenia barwnego ze względu na rozkład czopków na siatkówce
Skoro różne rodzaje czopków są tak nierównomiernie rozłożone na powierzchni siatkówki, warto uświadomić sobie, jaki ma to związek z zakresem widzenia barwnego. Zacznijmy od ustalenia, jak wiele czopków znajduje się w różnych częściach siatkówki ludzkiego oka. Od ponad 30 lat tym zagadnieniem zajmuje się Christine A. Curcio ze swoim zespołem (1987; 1990).
Na ryc. 14 w rozdziale poświęconym systemowi analizy zawartości obrazu przedstawiłem schematyczny rozkład czopków i pręcików w zależności od odległości od dołka centralnego. Z kolei na ryc. 114 w tym rozdziale można się zorientować, ile mniej więcej czopków znajduje się w różnych częściach siatkówki. Już na pierwszy rzut oka widać, że dysproporcje są ogromne: od 199 tys. na mm2 w samym środku dołka centralnego do 2 tys. na peryferiach siatkówki. Ale również na pozostałej powierzchni częstość występowania czopków jest bardzo zróżnicowana.
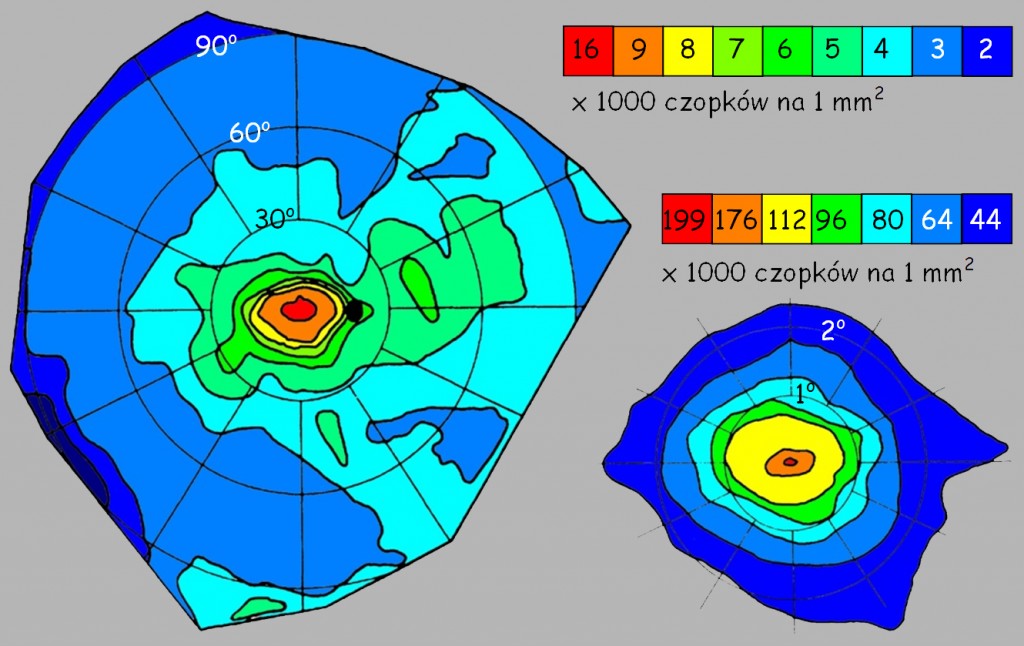
Ilościowy rozkład czopków, przedstawiony na ryc. 114 mówi nam coś o rozdzielczości, która przekłada się na wyrazistość widzenia w dobrych warunkach oświetleniowych. Nie mówi natomiast nic o widzeniu barwnym, a przecież czopki R, G i B także rozkładają się nierównomiernie na siatkówce. Powoduje to pewne problemy z trafnym rozpoznawaniem poszczególnych barw w zależności od tego, w jakiej odległości od dołka centralnego rzutowana jest na siatkówkę fala świetlna o określonej długości. Zależności te dla prawego oka przedstawia ryc. 115 (obraz dla lewego oka jest symetryczny).
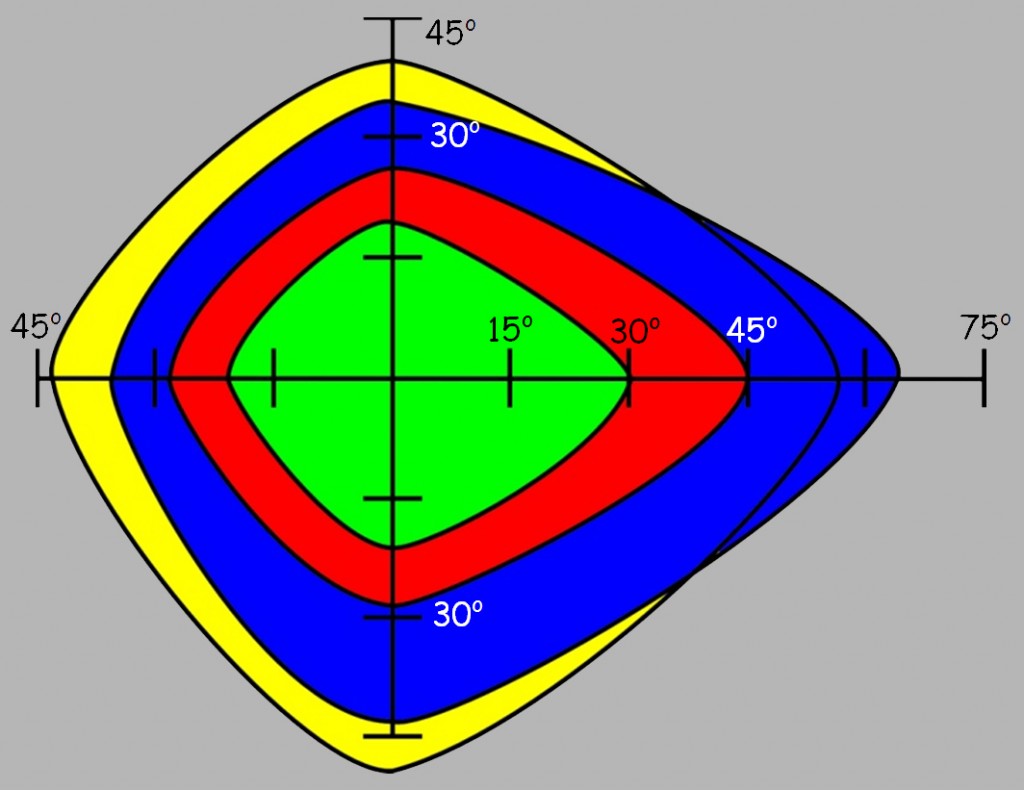
Przede wszystkim okazuje się, że w odległości większej niż 45o od dołka centralnego rozpoznawanie niemal wszystkich barw jest znacznie ograniczone. Możemy również stwierdzić, że barwa niebieska i żółta są trafniej rozpoznawane w większym zakresie niż barwa zielona i czerwona. Granice widzialności poszczególnych barw są nieco szersze. Eileen G. Ancman (1991) ustaliła, że powyżej 83,1o w zasadzie nie widać już barwy niebieskiej, powyżej 76,3o niknie barwa czerwona, a powyżej 74,3o zupełnie znika wrażenie zieleni. Tym niemniej problemy z ich trafnym rozpoznawaniem zaczynają się już znacznie wcześniej.
Spróbujmy przełożyć te dane na sytuację oglądania dużego obrazu, np. Bitwy pod Grunwaldem, Jana Matejki (ryc. 116). Obraz ma rzeczywiście imponujące rozmiary: 4,26 m wysokości i 9, 87 m szerokości. Stosując wzory, które wykorzystaliśmy do szacowania zakresu ostrego widzenia bardzo łatwo możemy ustalić, ile centymetrów przypada na 15o, 30o, 45o i 60o kąta pola widzenia, jeżeli obraz będziemy oglądali z odległości, np. 5 m. I tak, 15o odpowiada ok. 1,3 m [2 x tg (15o / 2) x 5 m], 30o – 2,7 m, 45o – 4,1 m i 60o – 5,8 m. Teraz pozostaje już tylko przenieść te miary na obraz i ze zdziwieniem odkryć kilka prawidłowości.

Przypuśćmy, że przyglądamy się twarzy pochwyconego na powróz Markwarda von Salzbacha, komtura brandenburskiego z prawej strony obrazu, mniej więcej w odległości 9 m od jego lewej krawędzi. Zapytani znienacka o kolor żupana, w jaki ubrany jest książę Witold (ok. 5,5 m od lewej) lub o barwę kaptura wojownika stojącego z toporem przez wielkim mistrzem krzyżackim (ok. 3,5 m od lewej), możemy mieć z tym poważny problem, bowiem znajdują się oni na granicy trafnej rozpoznawalności barwy czerwonej. Z kolei przenosząc wzrok na lewą stronę obrazu, gdzie na tle rozwianych, czerwonych szat widać postać Jakuba Skarbka z Góry, atakującego Kazimierza V, księcia szczecińskiego, przedstawionego w hełmie z potężnym pióropuszem, raczej nie rozpoznamy poprawnie nie tylko barwy żupana księcia Witolda, ale tym bardziej zielonych szat komtura tucholskiego, Heinricha von Schwelborna, ginącego od miecza czeskiego rycerza Jana Žižki (ok. 7,5 m).
Przy okazji warto przypomnieć, że Jan Matejko od urodzenia miał znaczną wadę wzroku, co praktycznie uniemożliwiało mu widzenie swoich dzieł w całości. Oglądał je podczas malowania wyłącznie z perspektywy kilkunastu centymetrów, co znacznie zawężało jego pole rozpoznawania barw! Swoje obrazy oglądał w całości tylko w wyobraźni.
Teoria procesów przeciwstawnych Ewalda Heringa
Po krótkiej wizycie w Muzeum Narodowym w Warszawie pozostańmy nadal w roku 1878, w którym Jan Matejko ukończył Bitwę pod Grunwaldem. Wtedy to Ewald Hering, niemiecki fizjolog opublikował teorię widzenia barwnego, opartą na procesach przeciwstawnych (opponent process theory). Była to koncepcja alternatywna w stosunku do „obowiązującej” wówczas trójchromatycznej teorii widzenia barw Younga i Helmholtza. Podstawą swojej teorii Hering uczynił także wyniki badań Newtona nad rozszczepieniem światła białego na różnobarwne składowe. Odrzucił jednak hipotezę o istnieniu triady receptorów wrażliwych na barwę czerwoną, zieloną i niebieską twierdząc, że ta idea nie tłumaczy subiektywnego wrażenia barwy żółtej”, równie czystej i pierwotnej, jak czerwień, zieleń i błękit.
Hering zauważył, że w wyniku mieszania niektórych barw w różnych proporcjach powstają nowe odcienie, które stanowią tonalnie przejścia między nimi, a mieszanie innych nie daje takich efektów. Na przykład, odcienie barwy pomarańczowej są formą przejściową między barwą czerwoną a żółtą. Podobnie, jak odcienie purpury w różnym stopniu są nasycone czerwienią i błękitem. Są jednak również i takie zestawienia barw, które nie tworzą form przejściowych, lecz zupełnie nowe jakości. Na przykład, zsumowanie barwy żółtej i niebieskiej tworzy zupełnie nową jakość, czyli zieleń. Z kolei połączenie zieleni z czerwienią daje barwę błota. Na podstawie rozważań dotyczących subiektywnych doświadczeń widzenia barw Hering doszedł do przekonania, że w siatkówce oka powinny znajdować się komórki, które tworzą pary odpowiedzialne za percepcję opozycyjnych barw: czerwonej i zielonej oraz niebieskiej i żółtej, czyli tych, które mieszając się ze sobą tworzą nowe jakości, a nie formy pośrednie.
Formułując swoją koncepcję Hering zwrócił uwagę na podejmowane przez Newtona próby geometryzacji przestrzeni barw, które zaowocowały opisaniem przez niego całego spektrum światła widzialnego w postaci koła barw (ryc. 117A). Young i Helmholtz zignorowali ten element koncepcji Newtona i poprzestali na linearnym opisie sekwencji barw, zgodnie rosnącą długością odpowiadających im fal świetlnych (por. ryc. 110).
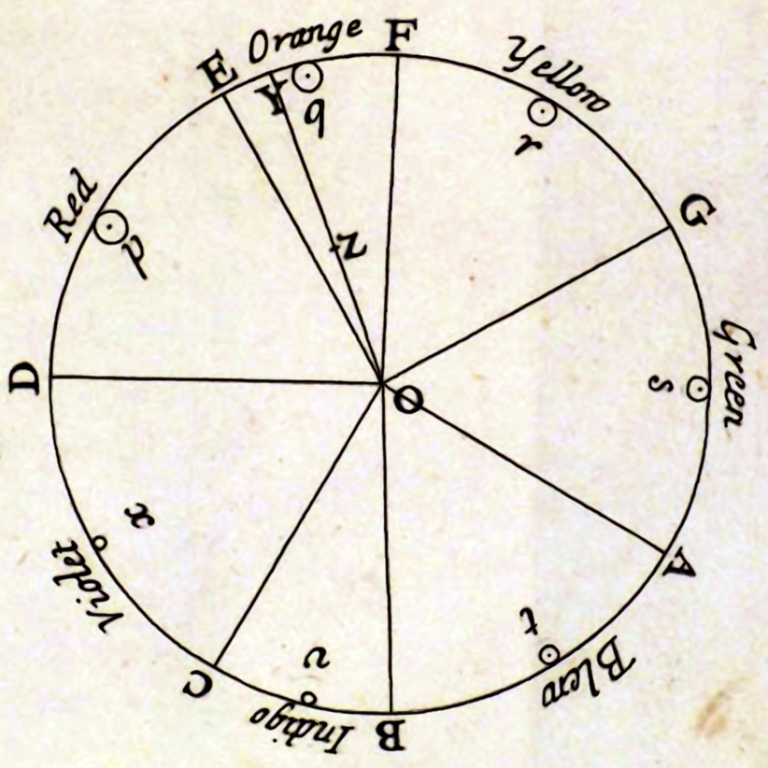
Z kolei Heringowi bardzo przypadł do gustu pomysł przedstawienia barw w formie koła, bowiem dostrzegł on, że ich rozkład znakomicie odpowiada dostrzeżonym przez niego opozycjom barwy czerwonej i zielonej oraz żółtej i niebieskiej. Leżąc po przeciwnych stronach koła, barwy opozycyjne nie tworzą ze sobą form przejściowych; tworzą je natomiast z barwami należącymi do drugiej pary. newtonowski rozkład barw na kole, Hering przedstawił w nieco bardziej schematycznej formie sugerując, że cztery podstawowe barwy: czerwień, żółcień, zieleń i błękit dzielą koło barw na cztery części, zajmując w ramach każdej z nich dominującą pozycję (ryc. 117B).
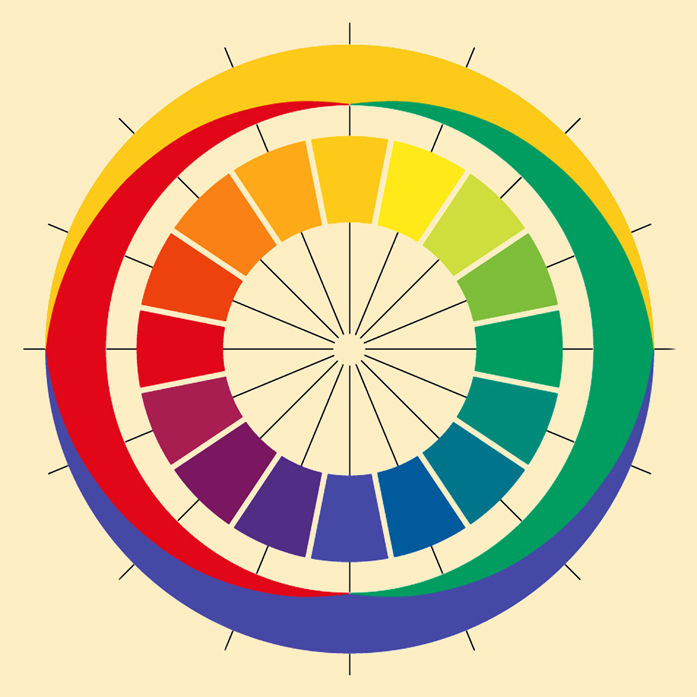
Gdyby Ewald Hering dożył 128 lat mógłby poczuć się w pełni usatysfakcjonowany po przeczytaniu artykułu Rogera N. Sheparda (1962), poświęconego próbie empirycznego odtworzenia umysłowej struktury podobieństwa 14 barw dobranych w zakresie długości fali świetlnej od 434 nm do 674 nm przez Göstę Ekmana (1954). Zadaniem osób badanych w eksperymencie Sheparda było porównanie ze sobą wszystkich tych barw w parach. Okazało się, że z psychometrycznego punktu widzenia rozwiązanie dwuwymiarowe jest w pełni satysfakcjonujące i niemal dokładnie odtwarza newtonowską strukturę relacji między barwami. I co najważniejsze, wynik badania potwierdził także sugerowane przez Heringa opozycje barw (ryc. 118).
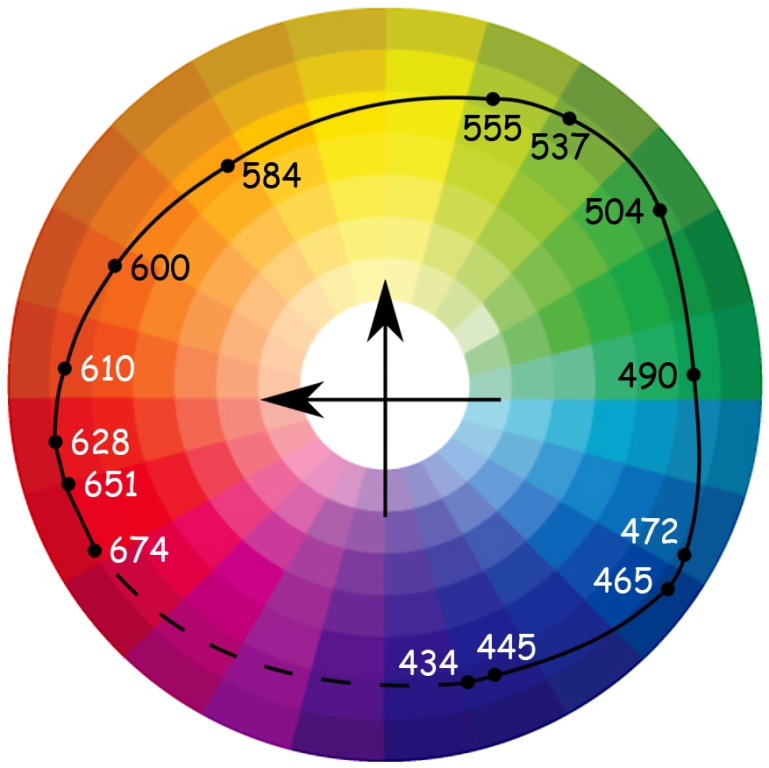
Roger N. Shepard i Lynn A. Cooper (1992) raz jeszcze przeprowadzili analogiczne badania w grupie osób normalnie widzących oraz w grupach osób cierpiących na deuteranopię (nie rozpoznają barwy zielonej) i protanopię (nie rozpoznają barwy czerwonej), a także nie widzących od urodzenia. Zmniejszyli także liczbę porównywanych barw do dziewięciu oraz prezentowali bodźce zarówno w formie wizualnej (barwne kartoniki), jak i w formie nazw. Okazało się, że rozkład barw prezentowanych za pomocą nazw był identyczny we wszystkich grupach, z wyjątkiem osób, które były niewidome od urodzenia. Istotne różnice między grupami wystąpiły natomiast wtedy, gdy barwy prezentowano w formie wizualnej.
Innym wsparciem koncepcji kolorów opozycyjnych Heringa jest doświadczenie powidoku barwnego (afterimage), zwanego również kontrastem następczym (ryc. 119). Jest to rodzaj iluzji wzrokowej, która powstaje na skutek długotrwałej stymulacji różnych części siatkówki różnymi barwami.
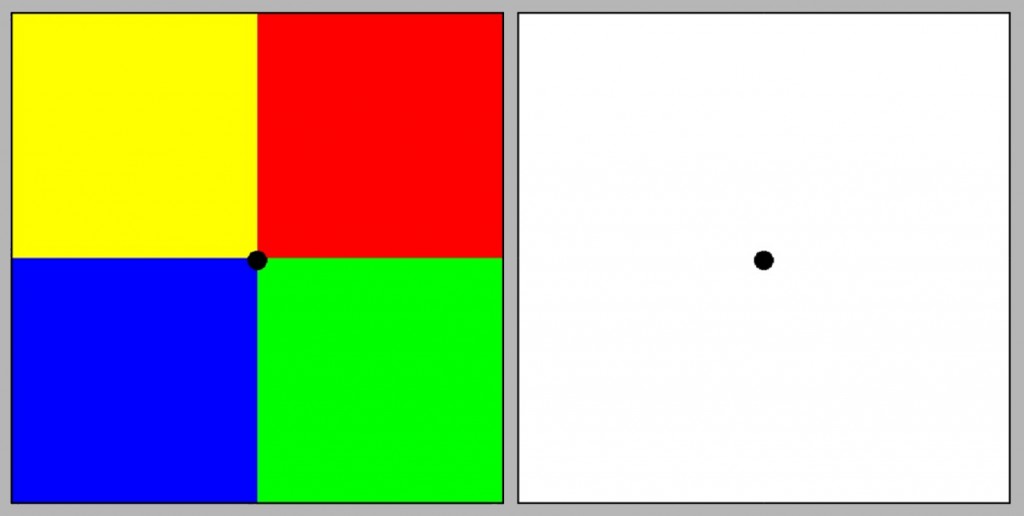
Żeby wywołać powidok należy przez 30 sekund skupić wzrok na czarnym punkcie leżącym na styku czterech barwnych kwadratów z lewej strony. Następnie należy przenieść wzrok na panel z prawej strony i także skupić wzrok na środkowym punkcie. Nieoczekiwanie na białej płaszczyźnie pojawią się wówczas cztery kwadraty w barwach dopełniających, tzn. w miejscu żółtego dostrzeżemy kolor zbliżony do niebieskiego, w miejscu czerwonego – kolor zielony itd. Zdaniem Heringa fakt, że powidoki przyjmują barwy opozycyjne jest kolejnym wsparciem dla jego koncepcji widzenia barwnego opartego na procesach przeciwstawnych.
Oprócz postulowania dwóch mechanizmów leżących u podłoża widzenia barw opozycyjnych, Hering zwrócił także uwagę na trzecią opozycję, tym razem w zakresie luminancji, czyli jasności oświetlenia. Twierdził, że utrata zdolności do barwnego widzenia, np. w warunkach obniżonego oświetlenia jest niezależna od zdolności do rozróżniania stopni jasności obrazu, w zakresie od bieli do czerni. Zdaniem Heringa, za kodowanie jasności musi być zatem odpowiedzialny trzeci typ receptora, a w związku z tym również trzeci przeciwstawny mechanizm, niezależny od tych, które leżą u podstaw widzenia opozycji barwnych.
Wszystkie trzy mechanizmy percepcyjne Hering ulokował w receptorach, znajdujących się w siatkówce oka. Odrzuciwszy, postulowany przez Younga i Helmholtza podział receptorów na trzy rodzaje, wyspecjalizowane w rejestrowaniu barwy czerwonej, zielonej i niebieskiej, sam również zaproponował trójpodział receptorów, ale według zupełnie nowych kryteriów. Przyjął bowiem, że w siatkówce są receptory odpowiedzialne za kodowanie barwy czerwonej i zielonej, żółtej i niebieskiej oraz bieli i czerni (ryc. 120).
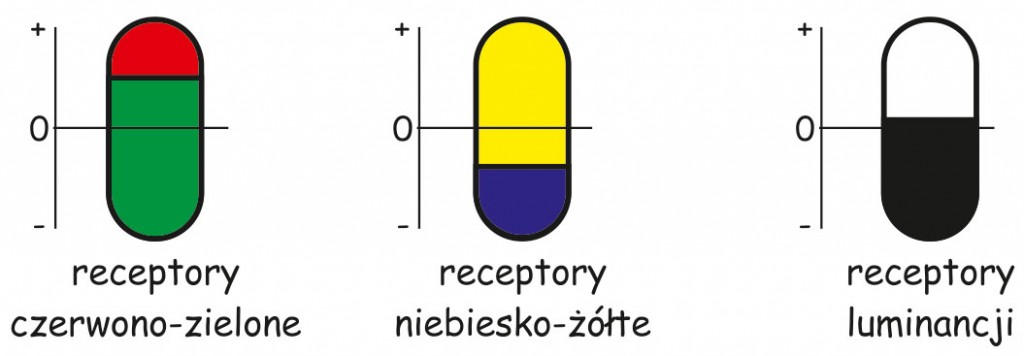
Jego zdaniem, zasada działania wszystkich typów receptorów opiera się na odbudowywaniu lub rozpadzie tej samej substancji chemicznej, znajdującej się wewnątrz nich. Na przykład, wzrost substancji w receptorach czerwono-zielonych, przy jednoczesnym wyzerowaniu jej stanu w receptorach niebiesko-żółtych i biało-czarnych byłby podstawą wrażenia czerwieni. Jeśli towarzyszyłby temu równocześnie rozpad, czyli zmniejszanie się ilości substancji chemicznej w receptorach niebiesko-żółtych, wówczas mielibyśmy wrażenie fioletu lub purpury, itd. Bezpośrednią przyczyną rozpadu lub gromadzenia się substancji chemicznej w poszczególnych receptorach jest – zdaniem Heringa – wpadające do oka światło o określonej długości.
Teoria stref widzenia barwnego Müllera-Hurvicha-Jameson
Do lat 30. XX wieku, trójchromatyczna teoria Younga-Helmholtza i teoria procesów przeciwstawnych Heringa były traktowane, jako dwie sprzeczne ze sobą koncepcje wyjaśniające mechanizm widzenia barw. Rozwiązanie tego teoretycznego impasu zaproponował George Elias Müller (1930 – za: Klein, 2010). Przedstawił on koncepcję stref kodowania barw na szlaku wzrokowym (zone theory). Zgodnie z jego sugestią, pierwsza strefa jest związana z reaktywnością siatkówkowych fotoreceptorów wrażliwych na światło czerwone, zielone i niebieskie. Strefa druga to droga wzrokowa na odcinku siatkówka oka – kora mózgu i trzecia, związana jest z aktywnością struktur korowych. Intuicje Müllera okazały się niezwykle trafne.
Pierwsza strefa, obejmująca struktury wewnątrz siatkówki oka, a w tym również receptory typu R, G i B została całkiem przyzwoicie opisana przez Younga i Helmholtza w ramach trójchromatycznej teorii widzenia barw. Druga strefa jest związana ze strukturą powiązań i aktywnością komórek zwojowych, wyprowadzających sygnał z siatkówki oka do LGN i dalej do korowych struktur mózgu. Dane płynące z fotoreceptorów są na tym odcinku porządkowane dokładnie w taki sposób, jak opisał to Hering w ramach swojej teorii procesów przeciwstawnych. Wreszcie trzecia strefa kodowania barw, obejmująca korowe obszary mózgu, począwszy od obszarów plamkowych w V1, a kończąc na strukturze V4 w płatach skroniowych, która została opisana przez Semira Zeki’ego (1973; 1993; 1999; 2003), a także przez Edwina H. Landa i Johna J. McCanna (1971) w teorii stałości widzenia barw retinex.
Koncepcja stref widzenia barwnego zaproponowana przez Müllera w latach 30. XX wieku, została w znacznym stopniu potwierdzona empirycznie w badaniach fizjologicznych i neurologicznych dopiero trzydzieści lat później. Zanim się to jednak stało, Leo Hurvich i Dorthea Jameson (1957), opierając się na pomysłach Müllera, przeprowadzili badania empiryczne, których celem było zweryfikowanie teorii procesów opozycyjnych poprzez ustalenie zależności między spostrzeganymi barwami a odpowiadającymi im długościami fali światła widzialnego.
Hurvich i Jameson podjęli próbę ilościowego opisania przebiegu procesów przeciwstawnych za pomocą metod psychofizycznych. Za Müllerem przyjęli, że w drugiej strefie szlaku wzrokowego dane pochodzące z trzech rodzajów fotoreceptorów wrażliwych na światło czerwone, zielone i niebieskie, generują wrażenia w zakresie czterech barw podstawowych, które wykluczają się wzajemnie w parach (czerwona vs zielona i niebieska vs żółta). Jeżeli tak jest, to w pełnym zakresie światła widzialnego można empirycznie ustalić takie długości fali elektromagnetycznej, które odpowiadają zarówno maksymalnym, jak i minimalnym wartościom nasycenia barw w obu parach. Rozumując analogicznie, można również wyznaczyć długości fali, które odpowiadają za różne stopnie jasności na wymiarze: biel vs czerń.
Osoby biorące udział w eksperymentach przeprowadzonych przez Hurvicha i Jameson dokonywały serii ocen dotyczących zawartości danej barwy, np. niebieskiej, we wszystkich barwach z całego spektrum światła widzialnego. Analogiczne zadanie dotyczyło pozostałych trzech barw: żółtej, czerwonej i zielonej.
Na podstawie zgromadzonych danych Hurvich i Jameson opisali nie tylko zależności między długością fali świetlnej a spostrzeganą intensywnością każdej z czterech barw, ale przede wszystkim wyznaczyli te długości fali elektromagnetycznej, które odpowiadały zerowemu nasileniu intensywności barw opozycyjnych w parach. Podobnie została opisana zależność między długością fali świetlnej a spostrzeganą jasnością światła (zob. ryc. 121).
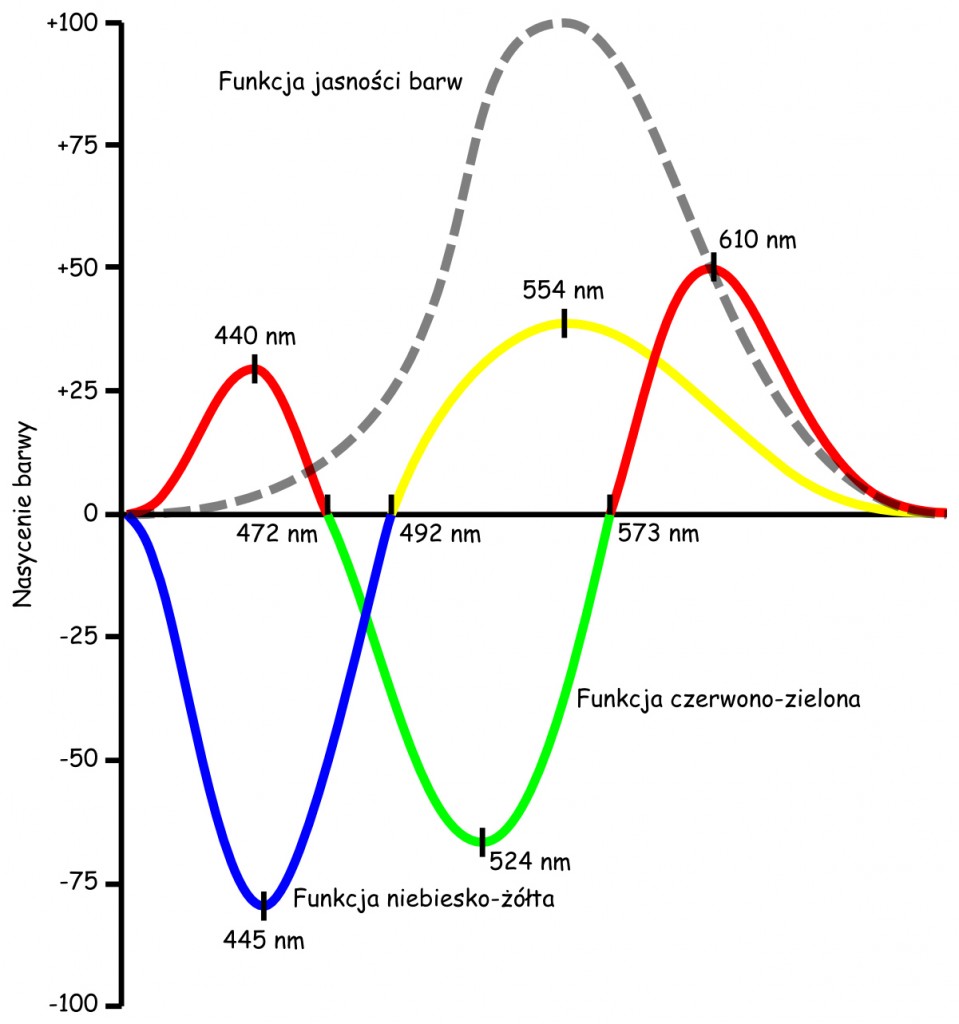
Na podstawie empirycznie ustalonych funkcji wykreślonych na ryc. 121 widać, że cztery podstawowe barwy są spostrzegane w dwóch opozycjach: niebiesko-żółtej i czerwono-zielonej. Wrażenie czystej barwy niebieskiej generuje fala światła widzialnego o długości 445 nm, a fala o długości 554 nm – barwy żółtej. Z kolei fala o długości 492 nm wywołuje wrażenie barwy zielonej, która jest całkowicie pozbawiona zarówno barwy żółtej, jak i niebieskiej. To sugerowany przez Heringa i Müllera neutralny punkt przejścia między tym dwiema opozycyjnymi barwami.
Nieco bardziej złożona jest sytuacja w odniesieniu do opozycji barwy czerwonej i zielonej. Wrażenie czystej zieleni odpowiada fali o długości 524 nm, natomiast wrażenie czystej czerwieni jest składową dwóch barw: czerwonej, o długości światła widzialnego 610 nm i fioletowej, o długości 440 nm. Barwy całkowicie pozbawione czerwieni i zieleni, czyli barwy neutralne dla tej opozycji, to błękit (472 nm) i żółcień (573 nm).
Ostatnia z funkcji przedstawionych na ryc. 121 to funkcja jasności. Na podstawie przebiegu tej funkcji możemy stwierdzić, że maksymalna jasność jest związana z długością fali świetlnej odpowiadającej czystej barwie żółtej i zmniejsza się symetrycznie wraz z wydłużaniem się i skracaniem długości fali elektromagnetycznej w kierunku krańców zakresu światła widzialnego.
Wyniki badań przeprowadzonych przez Hurvicha i Jameson potwierdziły teorię procesów przeciwstawnych Heringa w badaniach psychofizycznych. Do wyjaśnienia pozostała jeszcze kwestia neurofizjologicznych podstaw tych procesów, czyli odpowiedź na pytanie, jak to się dzieje, że na podstawie danych pochodzących z trzech receptorów wrażliwych na światło czerwone, zielone i niebieskie powstaje wrażenie widzenia barwy żółtej oraz wrażenia różnych stopni jasności. Na tę teorię nie trzeba było już długo czekać.
Od pobudzenia czopków R, G i B do widzenia barw opozycyjnych
Odkrycie związku między wrażliwością trzech rodzajów czopków w siatkówce oka na długość fali światła widzialnego a widzeniem barw opozycyjnych, opisanym w ramach teorii procesów przeciwstawnych, dokonano na podstawie wyników badań nad reakcjami komórek zwojowych, które odprowadzają sygnał z siatkówki oka do ciała kolankowatego bocznego (LGN). Jak pamiętamy, LGN znajduje się mniej więcej w połowie drogi między siatkówkami oczu a korą mózgu.
Najpierw Edward F. MacNichol i Gunnar Svaetichin (1958), prowadząc badania nad złotą rybką, a kilka lat później, Russell L. de Valois (1960) w analogicznych badaniach, prowadzonych na małpach stwierdzili, że aktywność niektórych komórek zwojowych w LGN znacznie wzrasta, gdy na siatkówkę oczu pada światło o długości odpowiadającej barwie, np. czerwonej, natomiast reakcja tych komórek zmniejsza się pod wpływem światła zielonego. Z kolei inne komórki reagują odwrotnie: światło zielone wyraźnie je pobudza, ale pod wpływem światła czerwonego ich aktywność spada. Co więcej, odkryto również komórki zwojowe, które intensywnie reagują na światło niebieskie, a ich aktywność wyraźnie się zmniejsza, gdy barwa światła zmienia się na żółtą. I wreszcie są takie, które intensywnie reagują na światło żółte natomiast ich aktywność jest wyhamowywana pod wpływem światła niebieskiego. W badaniach tych zidentyfikowano również komórki, które reagują na światło jasne a nie reagują na ciemne i komórki, które aktywizują się pod wypływem niewielkiego oświetlenia, a nie reagują na jasne świtało.
Reakcje wszystkich wymienionych typów komórek zwojowych Stephen E. Palmer (1999) określa, jako neuronalną implementację teorii procesów przeciwstawnych Heringa. Podstawą działania tych komórek jest opisany już mechanizm aktywności komórek dwubiegunowych i zwojowych tworzących tzw. kanały włączeniowe (typu ON) i wyłączeniowe (typu OFF). Przypomnę, że na przeważającej powierzchni siatkówki fotoreceptory są ze sobą połączone w koliste pola recepcyjne komórek horyzontalnych. Fotoreceptory znajdujące się w środkowych częściach tych pól dodatkowo łączą się z komórkami dwubiegunowymi typu ON lub OFF, a te z kolei, z komórkami zwojowymi także, odpowiednio, typu ON lub OFF. Mechanizm przekazywania danych dotyczących długości światła widzialnego w zakresie barw opozycyjnych za pośrednictwem tych kanałów jest analogiczny do mechanizmu kodowania jasności światła padającego na pole recepcyjne, opisanego w jednym z poprzednich rozdziałów.
Jeżeli pole recepcyjne komórki zwojowej, np. typu ON jest tak zorganizowane, że jego środkowa część reaguje na falę światła, odpowiadającą barwie zielonej, wówczas w momencie oświetlenia jej takim światłem do LGN, a następnie do V1 jest przekazywana informacja o obecności barwy zielonej, a hamowana jest informacja o obecności barwy czerwonej. Jeżeli natomiast środkowa część pola recepcyjnego komórki zwojowej jest typu OFF, wówczas informacja o oświetleniu tej części siatkówki światłem o długości odpowiadającej barwie zielonej jest wyhamowywania, a torowana jest informacja o obecności barwy czerwonej. Podobnie działają kanały włączeniowe i wyłączeniowe w odniesieniu do pary barw niebieskiej i żółtej (Daw, 2008).
Do wyjaśnienia pozostaje już tylko związek między reakcją komórek zwojowych kodujących jeden z czterech kolorów podstawowych a reaktywnością fotoreceptorów R, G i B. Zagadnienie to nie jest ostatecznie rozstrzygnięte do dzisiaj, ale Russell L. de Valois, Israel Abramov i Gerald H. Jacobs (1966) zaproponowali rozwiązanie, które nie zostało dotąd podważone. Jego graficzna ilustracja znajduje się na ryc. 122.
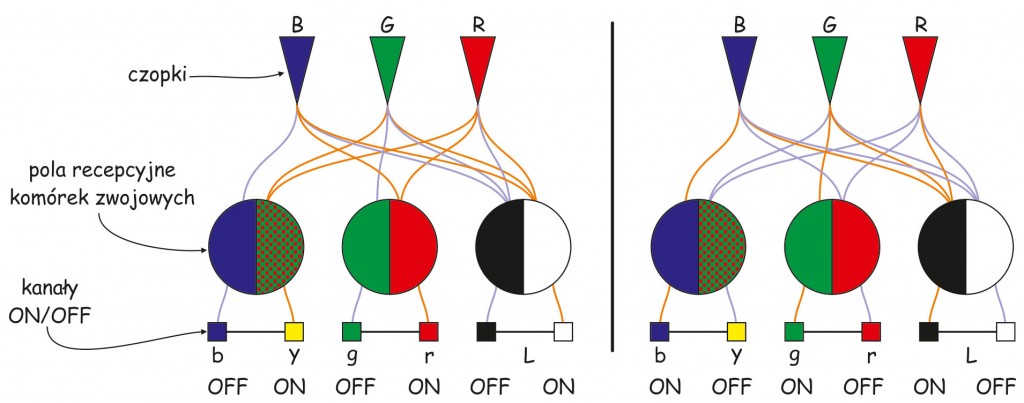
Zgodnie ze schematem przedstawionym na ryc. 122, wrażenie barwy czerwonej może powstać w wyniku:
- aktywizacji kanału ON dla pola recepcyjnego, które w środkowej części odbiera dane o poziomie pobudzenia czopków typu R i B lub
- aktywizacji kanału OFF dla pola recepcyjnego, które z otoczki odbiera dane o poziomie pobudzenia czopków typu R i B.
Z kolei wrażenie barwy zielonej może powstać w wyniku:
- aktywizacji kanału ON dla pola recepcyjnego, które w środkowej części odbiera dane o poziomie pobudzenia czopków typu G lub
- aktywizacji kanału OFF dla pola recepcyjnego, które z otoczki odbiera dane o poziomie pobudzenia czopków typu G.
Wrażenie barwy żółtej także powstaje w jednej z dwóch sytuacji:
- aktywizacji kanału ON dla pola recepcyjnego, które w środkowej części odbiera dane o poziomie pobudzenia czopków typu R i G lub
- aktywizacji kanału OFF dla pola recepcyjnego, które z otoczki odbiera dane o poziomie pobudzenia czopków typu R i G.
Również wrażenie barwy niebieskiej może powstać w dwóch sytuacjach:
- aktywizacji kanału ON dla pola recepcyjnego, które w środkowej części odbiera dane o poziomie pobudzenia czopków typu B lub
- aktywizacji kanału OFF dla pola recepcyjnego, które z otoczki odbiera dane o poziomie pobudzenia czopków typu B.
Oczywiście wrażenie którejkolwiek barwy powstaje tylko wtedy, gdy wszystkie podsystemy odpowiedzialne za przetwarzanie danych sensorycznych, na każdym etapie szlaku wzrokowego działają sprawnie. Jeśli nie, to finalne efekty przetwarzania danych mogą nawet znacznie odbiegać od opisanych w powyższych schematach.
I na koniec raz jeszcze przypomnienie, w jaki sposób działa mechanizm detekcji jasności (luminancji). Otóż wrażenie jasności może powstać w wyniku:
- aktywizacji kanału ON dla pola recepcyjnego, które w środkowej części odbiera dane o poziomie pobudzenia wszystkich rodzajów czopków lub
- aktywizacji kanału OFF dla pola recepcyjnego, które z otoczki odbiera dane o poziomie pobudzenia wszystkich rodzajów czopków.
Ze względu na różną jasność światła sprzężoną z różną długością fali elektromagnetycznej, znacznie większy wpływ na wrażenie jasności będzie miał poziom pobudzenia czopków R i G, niż B, ponieważ ich maksymalny poziom wrażliwości, odpowiednio 560 i 530 nm, pokrywa się z tą długością fali świetlnej, która jest najjaśniejsza, czyli ok. 550 nm (zob. wykres funkcji jasności na ryc. 121).
BARWY W OBRAZACH
W historii sztuki dzieje koloru i jego teorii to oscylowanie między degradacją do roli ozdoby czy ornamentu a awansem do podstawowej prawdy malarstwa (Melville, 1994)
O eksperymentach wizualnych Claude Moneta
Czy uzbrojeni w wiedzę dotyczącą widzenia barwnego potrafimy lepiej zrozumieć, dlaczego malarze lub fotograficy korzystają z kolorów w taki lub inny sposób i jakie osiągają dzięki temu efekty wizualne?
Pierwszy przykład obrazu, na jaki chciałbym zwrócić uwagę to Impresja. Wschód słońca, Claude Moneta (ryc. 123 A).

Oglądając reprodukcję tego obrazu w albumie, Margaret Livingstone (2002) zauważyła interesujące zjawisko. Niemal narzucająca się obserwatorowi krwistoczerwona kula wschodzącego słońca przestaje budzić zainteresowanie wtedy, gdy jest ono oglądane w wersji monochromatycznej (ryc. 123 B). Jej zdaniem usunięcie barw sprawia, że słońce jakby zanika, chociaż nie jest tak, że w wersji monochromatycznej jego kontury są zupełnie niewidoczne.
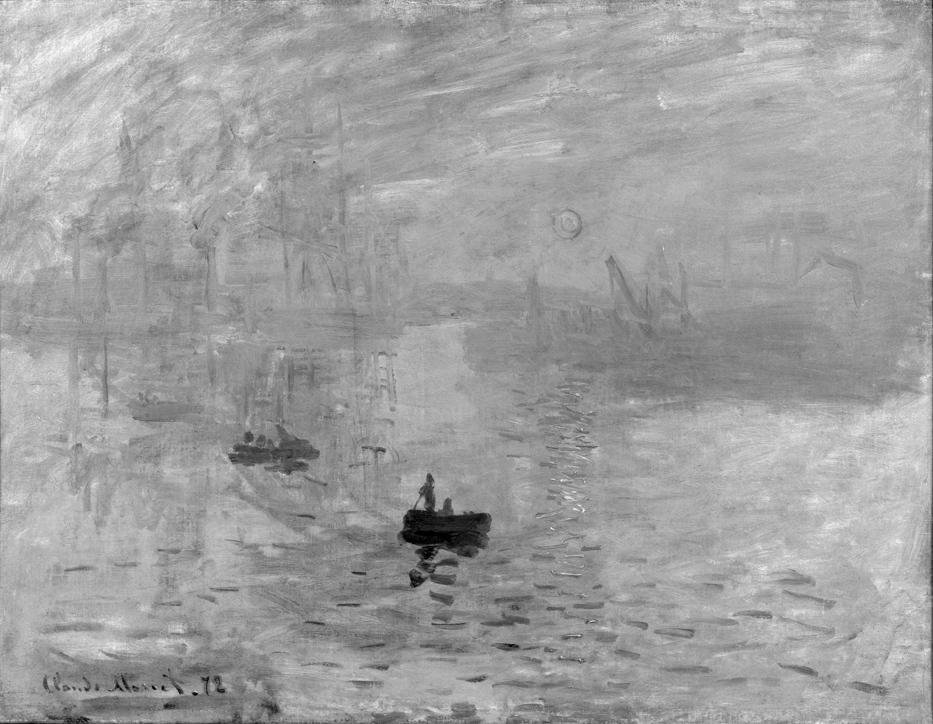
Żeby przekonać się, czy rzeczywiście obraz Moneta jest inaczej oglądany w wersji oryginalnej i monochromatycznej, razem z Anną Szpak i Patrycją Kot przeprowadziliśmy badanie, podczas którego barwną reprodukcję Impresji pokazaliśmy 19 kobietom i 19 mężczyznom, a jego wersję monochromatyczną – 26 kobietom i 17 mężczyznom. Wszystkie osoby były w wieku ok. 22 lat. Obrazy eksponowaliśmy na 23” monitorze Apple Cinema HD Display (1920 x 1200 pikseli). Podczas eksperymentu rejestrowaliśmy ruch gałek ocznych osób badanych za pomocą okulografu SMI iView X Hi Speed 1250 Hz.
Oczekiwaliśmy, że zainteresowanie okolicą słońca na obrazie barwnym będzie znacznie większe, niż zainteresowanie analogicznym obszarem na obrazie monochromatycznym. Zastanawialiśmy się także, jak wiele uwagi osoby badane poświęcą innym fragmentom obrazu, np. takim, jak łódź, znajdująca się na pierwszym planie, która w obu wersjach obrazu jest bardzo wyraźna. Zwłaszcza w odniesieniu do wersji monochromatycznej przypuszczaliśmy, że to właśnie ciemna łódź z pierwszego planu, która silnie kontrastuje z jasnym tłem morza, powinna wzbudzić znacznie większe zainteresowanie osób badanych niż okolica słońca.
Zapis okulograficzny w pełni potwierdził obserwacje Livingstone i nasze oczekiwania (ryc. 123 C i D). Podczas oglądania barwnej reprodukcji osoby badane zdecydowanie najwięcej czasu poświęciły na przyglądanie się słońcu (ryc. C), natomiast oglądając reprodukcję monochromatyczną szczególnie interesowała ich łódź na pierwszym planie (ryc. D). Wszystkie różnice w czasach fiksacji wzroku na tych samych fragmentach obu wersji obrazu oraz w porównaniu do innych fragmentów w ramach tych samych wersji są wyraźne i istotne statystycznie (HSD Tukeya; p < 0,001).


Jeżeli przyjmiemy, że czas poświęcany na oglądanie jakiegoś obiektu wiąże się z zainteresowaniem nim, to warto zastanowić się, co wzbudziło tak duże zaciekawienie osób badanych w tej ostro zarysowanej, czerwonej kuli na tle porannego nieba. Jako ciekawostkę można dodać, że oglądając obraz w muzeum i wpatrując się dłużej w słońce niektórzy doświadczają czegoś w rodzaju pulsowania, co prawdopodobnie jeszcze bardziej ich zaciekawia i zatrzymuje wzrok na tym miejscu.
Zdaniem Livingstone (2002) owo dziwaczne (weird) wrażenie bierze się stąd, że system wzrokowy przetwarzając dane dotyczące barw oraz jasności obrazu niezależnie od siebie nie może podjąć najbardziej fundamentalnej decyzji dotyczącej obecności vs nieobecności rzeczy w scenie wizualnej. Znacznie starsza ewolucyjnie część systemu wzrokowego odpowiedzialna za detekcję luminancji jest bezradna wobec zasadniczego braku kontrastów między sąsiadującymi ze sobą płaszczyznami słońca i nieba. Z kolei, ewolucyjnie młodsza część systemu wzrokowego, która jest wrażliwa na barwy, rejestruje na obrazie obecność czerwonego okręgu na tle błękitnego nieba. W rezultacie powstaje niesamowite (eerie) doświadczenie pulsowania, które jest reakcją systemu detekcji luminancji na trudność w określeniu pozycji obiektu, którego obecność jest zgłaszana przez system detekcji barwnej. W ten sposób można wyjaśnić efekt niestabilności fragmentu obrazu zbudowanego z dwóch sąsiadujących ze sobą barw o tej samej jasności. Krótko mówiąc, jasność słońca jest niemal taka sama, jak jasność tła, na którym umieścił go „nieświadomy neurobiolog”, Claude Monet. I w tym tkwi magia tego obrazu.
Łatwo można się przekonać, że jasność słońca i otaczającego go nieba jest bardzo podobna. Na ryc. 124 A znajduje się powiększony fragment obrazu Moneta, przedstawiający interesujące nas słońce. Poniżej jest ten sam fragment w wersji monochromatycznej (ryc. 124 B). Oszacowałem i naniosłem nań wartości liczbowe odpowiadające jasności obszarów leżących wokół nich. Jak widać, różnice między jasnościami wewnątrz i na zewnątrz słońca są niewielkie. Warto przy tym pamiętać, że niektóre ciemniejsze miejsca wewnątrz i na krawędziach słońca nie powstały z powodu użycia ciemniejszej barwy czerwonej, ale są cieniem rzucanym przez grubo kładzioną farbę.


Analiza obrazu Moneta ujawnia jeszcze jedną bardzo ważną cechę systemu wzrokowego, który korzysta z dobrodziejstwa rejestracji świata w barwach. Skoro system luminancji obserwatora ma trudności ze zidentyfikowaniem słońca na obrazie Moneta, a mimo to widzi je, to może to znaczyć tylko tyle, że barwa, podobnie, jak światłocień, także koduje kształt rzeczy, na które patrzymy. Żeby się o tym przekonać, wyizolowałem z oryginalnej wersji obrazu Moneta dwa obrazy złożone tylko z opozycyjnych barw: czerwonej i zielonej oraz żółtej i niebieskiej, ustalając ich luminancję na takim samym, średnim poziomie. Zabieg ten pozwolił stwierdzić ile czerwieni, zieleni, błękitu i żółci zawiera się w barwach, którymi Monet posłużył się do jego namalowania. Powstałe w ten sposób obrazy umożliwiają dostrzeżenie granic między barwnymi plamami, a tym samym odczytanie kształtów, które one kodują. W każdym razie takie są przewidywania wynikające z teorii procesów przeciwstawnych, zbudowanej na trafnych intuicjach Ewalda Heringa. Efekt symulacji procesów przeciwstawnych jest przedstawiony na ryc. 125 A‑D.




Obie pary barw opozycyjnych na ryc. 125 A i C mają taką samą luminancję. Powoduje to, że granice barw są słabo widoczne, ale z pewnością mózg nie ma kłopotów z ich zróżnicowaniem, choćby z tego powodu, że są kodowane z pomocą różnych kanałów, zbudowanych z komórek zwojowych, które zajmują się przesyłaniem danych sensorycznych w głąb mózgu. Wyłącznie dla celów ilustracyjnych, opracowałem również obie wersje obrazu w wysoko kontrastowej wersji monochromatycznej (ryc. 125 B i D). Teraz już nie powinno być żadnych wątpliwości, że podstawą widzenia słońca na obrazie Moneta nie jest kontrast jasności, lecz kontrast opozycyjnych barw, które wyraziście kodują kształt słońca (por. ryc. 124 B vs. ryc. 125 B i D).
Warto również zwrócić uwagę na to, że podczas oglądania barwnej reprodukcji obrazu drugim, najdłużej oglądanym obiektem był ciemny zarys łodzi, znajdującej się na pierwszym planie. W kontekście tego, co napisałem na temat rozbieżności informacji płynącej z kanałów barwnych i kanału luminancji trudno jest rozstrzygnąć, dlaczego osoby badane poświęcały tyle czasu na przyglądanie się temu fragmentowi obrazu. Z jednej strony, nie ma wątpliwości co do tego, że ze względu na kontrast jasności jest to najwyraźniejszy obiekt w całym obrazie, co znakomicie widać na jego monochromatycznej wersji (zob. ryc. 123 B). Z tego punktu widzenia możemy stwierdzić, że kanał luminancji „pozwala” na jednoznaczne zidentyfikowanie przedmiotu w obrazie, ze względu na jego wysoką kontrastowość.
Z drugiej jednak strony, łódź po prostu znika, gdy obraz zostanie przefiltrowany przez oba kanały barw opozycyjnych (ryc. 125 A‑D). Czyżby dłuższy czas przyglądania się łodzi był kolejnym przejawem konfliktu wizualnego między kanałem kodującym kształt na podstawie różnic w luminacji oraz na podstawie zróżnicowania barw opozycyjnych, który podwyższa zaciekawienie obserwatorów? A może to tylko przejaw naturalnej reakcji systemu wzrokowego, wyrażającej się podwyższoną koncentracją uwagi na tych elementach sceny wizualnej, które najtrafniej pozwolą odtworzyć jej sens.
No cóż, z pewnością nie uda nam się od razu rozwiać wszystkich wątpliwości. Warto jednak podrążyć jeszcze nieco problem słońca i powodów szczególnego zainteresowania nim. Z pewnością, analizowanego obrazu Moneta nie można zaliczyć do zbioru w pełni kontrolowanych bodźców eksperymentalnych. Żeby wyciągać na jego podstawie wnioski naukowe potrzebny jest nieco bardziej precyzyjny model.
Słońce Moneta pod lupą
Istota tzw. efektu Moneta, że tak pozwolę sobie określić analizowane zjawisko, polega na wzmożonym zainteresowaniu obserwatora tymi obszarami sceny wizualnej, które charakteryzują się rozbieżnością między danymi kodującymi kształty znajdujących się tam obiektów na podstawie luminancji i dwóch kanałów barwnych. Do namalowania tego fragmentu Monet zastosował barwę czerwoną (słońce) i niebieską (niebo) o podobnej luminancji, równiej ok. 40 jednostek (por. ryc. 124). Wzorując się na tym pomyśle, wspólnie z Piotrem Szyjką, skonstruowaliśmy bodźce i przeprowadziliśmy eksperyment, w którym wykorzystaliśmy okulograf (Francuz, Szyjka, 2011).
Materiał bodźcowy składał się z 48 plansz. Na każdej planszy znajdowały się 24 różnobarwne gwiazdki o tej samej wielkości [32,3 mm; 125 pikseli]. Gwiazdki były rozmieszczone losowo na szarym tle o takiej samej jasności [Lab = 50:0:0; RGB = 119:119:119]. Każdej gwiazdce był przypisany losowo kąt rotacji wokół swojej osi. Odległość środków poszczególnych gwiazdek od siebie była równa lub większa, niż zakres pola widzenia za pomocą dołka centralnego, czyli co najmniej 52,41 mm; [201,57 pikseli]. Na połowie plansz wszystkie gwiazdki miały po 7 ramion, natomiast na drugiej połowie plansz jedna z dwudziestu czterech gwiazdek miała 8 ramion. Na każdej z tych plansz zawsze była to gwiazdka o innej barwie kolorze (zob. przykładową planszę zawierającą gwiazdkę ośmioramienną na ryc. 126 A).


Jasność i barwa poszczególnych gwiazdek były w pełni kontrolowane. Przyjęliśmy trzy poziomy jasności gwiazdek: L = 25 (ciemniejsze niż tło), L = 50, czyli tyle samo ile wynosiła jasność tła oraz L = 75 (jaśniejsze niż tło). Na każdej planszy znajdowało się po osiem gwiazdek jaśniejszych, niż tło, po 8 gwiazdek ciemniejszych, niż tło i po 8 gwiazdek o takiej samej jasności, jak tło. Na ryc. 126B ilustruję monochromatyczny efekt powstały po usunięciu barwy z planszy znajdującej się po lewej stronie.
Barwy gwiazdek powstały z kombinacji parametrów odpowiadających wartościom granicznym i zerowym na skalach barwnych: od czerwieni do zieleni i od niebieskiego do żółtego. W rezultacie uzyskaliśmy 8 różnicowanych barw, co w połączeniu z trzema poziomami luminacji dało 24 różnobarwne i różno-jasne gwiazdki, które umieściliśmy losowo na poszczególnych planszach.
Liczba plansz prezentowanych osobom badanym wynikała stąd, że chcieliśmy, aby każda barwa o każdym poziomie jasności występowała, co najmniej jeden raz w wersji 8‑ramiennej. Ponieważ zadaniem osób badanych było odpowiedzenie na pytanie, czy na planszy znajduje się gwiazdka 8‑ramienna, dlatego połowa plansz ją zawierała, a druga połowa – nie. Ostatecznie wszystkie plansze pokazywaliśmy każdej osobie badanej trzykrotnie, w kolejności losowej. Do prezentacji bodźców i gromadzenia danych o reakcjach osób badanych wykorzystaliśmy procedurę napisaną w programie e‑Prime.
Wszystkie plansze eksponowaliśmy na 23″ Apple Cinema HD Display (1920 by 1200 pixels). Osoby badane siedziały w odległości ok. 50 cm od monitora i reagowały za pomocą klawiatury Ergodex, o zmiennym układzie klawiszy.
Przebadaliśmy 23 osoby (13 kobiet i 11 mężczyzn) w wieku ok. 23 lat. Tak jak wspomniałem, ich zadaniem było (trzykrotne, w porządku losowym) obejrzenie każdej planszy i udzielenie odpowiedzi na pytanie, czy kształty wszystkich gwiazdek na planszy są takie same. Podczas badania rejestrowaliśmy ruch gałek ocznych osób badanych za pomocą okulografu SMI iView X Hi Speed 1250 Hz. Jeżeli osoba badana twierdziła, że wszystkie gwiazdki mają taki sam kształt, wówczas pojawiała się następna plansza. Jeśli natomiast twierdziła, że nie wszystkie gwiazdki mają taki sam kształt, wówczas była proszona o to, aby przez 5 sek. skupiała wzrok na tej gwiazdce, o której twierdziła, że jest inna. To był wskaźnik trafności jej odpowiedzi. Oprócz zachowań okoruchowych rejestrowaliśmy także czas reakcji.
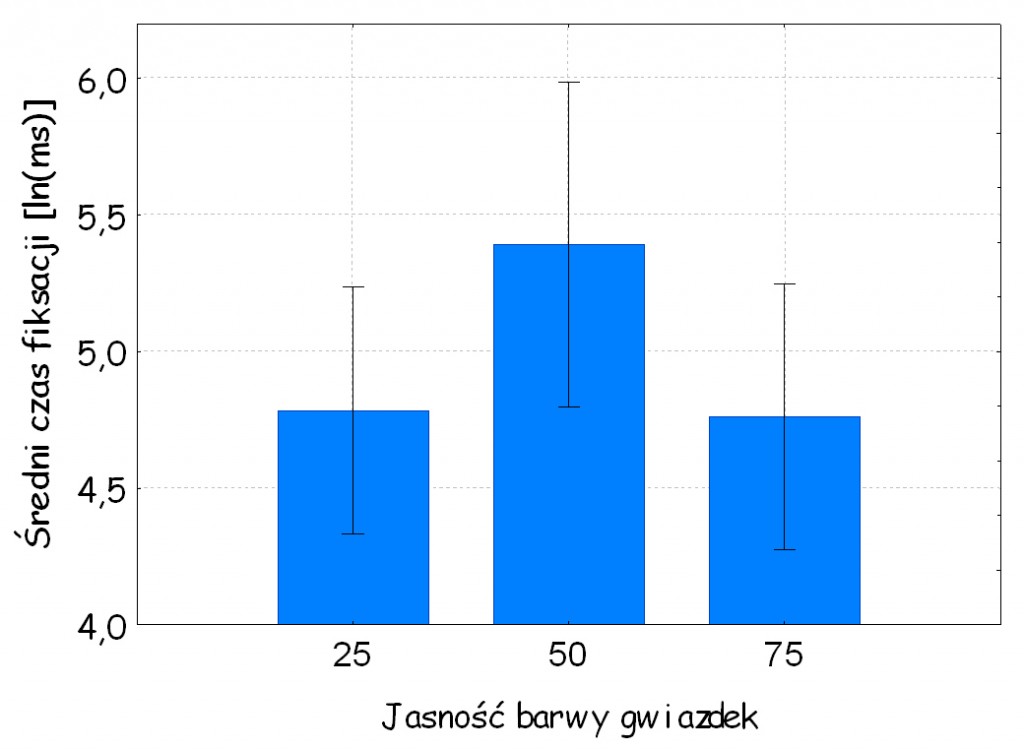
Chociaż w badaniu prosiliśmy osoby o to, aby aktywnie poszukiwały gwiazdek o odmiennych kształtach, tym niemniej przede wszystkim interesowało nas to, jak wiele uwagi poświecą one na przyglądanie się tym gwiazdkom, których luminancja była taka sama, jak jasność tła w porównaniu do tych, których luminancja była wyższa lub niższa od tła. Jeżeli efekt Moneta jest faktem, to czas poświęcony na przyglądanie się gwiazdkom o jasności równej jasności tła powinien być dłuższy niż czas przyglądania się pozostałym gwiazdkom.
Wyniki analizy wariancji w pełni potwierdziły te przypuszczenia. Nie tylko okazało się, że osoby badane dłużej przyglądały się gwiazdkom o jasności równej jasności tła, niż jaśniejszym lub ciemniejszym [F (2, 46) = 9,77; p < 0,001; η2 = 0,30; ryc. 127A], ale zależność ta potwierdziła się także w odniesieniu do gwiazdek 8‑ramiennych o luminancji równej 50 jednostek [F (2, 46) = 8,79; p < 0,001; η2 = 0,28]. Ponadto okazało się, że efekt Moneta występuje niezależnie od tego, jakiej barwy jest gwiazdka. Ważne, żeby jej jasność była taka sama, jak jasność tła.
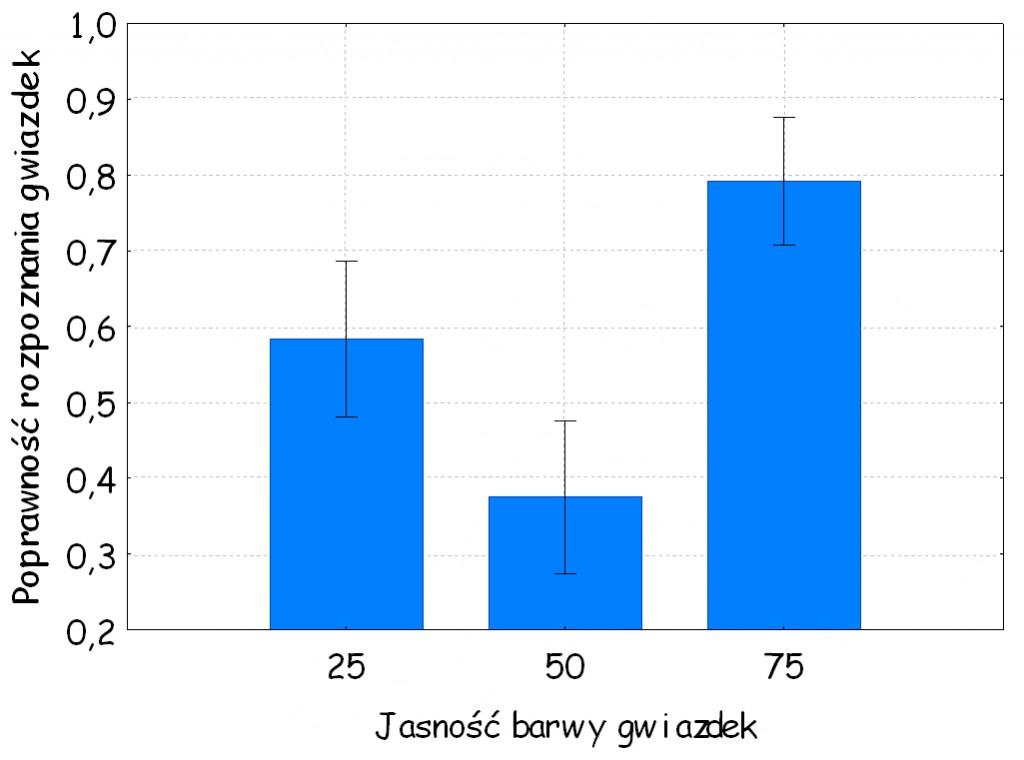
Dodatkowo stwierdziliśmy, że chociaż osoby badane poświęcały sporo uwagi gwiazdkom 8‑ramiennym o jasności równej jasności tła, tym niemniej, niezależnie od barwy gwiazdek, rzadziej poprawnie rozpoznawały je jako odmienne od pozostałych. W odniesieniu do niektórych barw różnice okazały się statystycznie istotne [np. w odniesieniu do barwy zielonej: F (2, 46) = 5,58, p = 0,007, η2 = 0,19; ryc. 127 B]. Wygląda to trochę tak, jakby rozwiązywanie problemu obecności gwiazdki na planszy ze względu na luminancję tak silnie dominowało ich uwagę, że nie byli w stanie poprawnie stwierdzić, że mają one po osiem, a nie po siedem ramion. Oto przykład, w jaki sposób umiejętne zwężenie pola uwagi obserwatora poprzez zadanie mu trudnego zadania percepcyjnego może zakłócić jego normalne procesy poznawcze. Kiedy gwiazdki są jasne i wyraźnie odcinają się od tła, wówczas prawdopodobieństwo ich trafnego rozpoznania wzrasta do 0,8, ale gdy ich jasność stanie się taka sama, jak jasność tła – trafność ich rozpoznania spada do niespełna 0,4.
Ile jest kształtów w jednej rzeczy?
W świetle przedstawionych wyników analiz obrazu Moneta warto rozważyć, jakie mogą być jeszcze inne konsekwencje rozbieżności między kształtem kodowanym za pośrednictwem kanału luminancji i kanałów barwnych?
Zacznijmy od przykładu obrazu Apollo i dwie Muzy, namalowanego w XVIII wieku przez Pompeo Girolamo Batoniego (ryc. 128 A). Obraz pochodzi z kolekcji Muzeum Pałac w Wilanowie w Warszawie. Przedstawia on trzy mitologiczne postaci: Apolla, symbol idealnej boskiej muzyki – harmonii sfer niebieskich oraz wpatrzone weń muzy – Eutrepe i Uranię, które uosabiają Muzykę i Astronomię (Gutowska-Dudek, 2005). To oczywiście jest perspektywa historyka sztuki. Natomiast laik w dziedzinie historii sztuki, oglądając ten obraz z pewnością dostrzeże młodego, roznegliżowanego młodzieńca w towarzystwie równie, przewiewnie odzianych dziewcząt, zaś „zatrzymane w kadrze” zachowanie się całej trójki może na nim sprawić wrażenie, co najmniej dwuznaczne (Francuz, 2012).
Niezależnie jednak od wiedzy obserwatora raczej nie mamy wątpliwości odnośnie do osób i rzeczy przedstawionych na obrazie. Odwołując się do klasycznych kanonów piękna możemy nawet uznać, że obraz jest bez zarzutu, ale też nie rodzi napięć, jakie zatrzymały nas dłużej przed obrazem Moneta. I nie ma w tym nic dziwnego, ponieważ system wzrokowy niewiele ma tutaj zagadek do rozwiązania. Kształty zakodowane zarówno za pomocą jasności, jak i dwóch kanałów barwnych (czerwono-zielonego i niebiesko-żółtego) potwierdzają się wzajemnie niemal w 100% (zob. ryc. 128 C‑D).




Żeby rozpoznać scenę przedstawioną na obrazie przez Batoniego, właściwie nie ma większego znaczenia, którą jego wersję oglądamy. W podobny sposób wszystkie trzy kanały wzrokowe kodują kształty namalowanych na nim postaci. Jest to najbardziej typowy sposób, w jaki system wzrokowy rejestruje i interpretuje rozkład światła wpadającego do oka obserwatora. Jest on najbliższy codziennemu doświadczeniu. W taki też sposób przez całe stulecia malarze zachodnioeuropejscy usiłowali uwiecznić rzeczywistość na obrazach. Dopiero wynalezienie fotografii w pierwszej połowie XIX wieku zdecydowanie załamało ten kierunek rozwoju malarstwa. W rezultacie sztuki wizualne uwolniły się ze zbyt ciasnych ram epoki stylów i prawdziwie eksplodowały rewolucyjnymi zmianami w ramach tzw. sztuki współczesnej.
Obraz Claude Moneta, Impresja. Wschód słońca, jest jednym z najważniejszych przejawów tej rewolucji. Monet zerwał z tradycją pokazując, że celem sztuki wizualnej nie jest wierne przedstawianie rzeczywistości, ale przeciwnie, jej kreowanie. W gruncie rzeczy każdy intencjonalnie stworzony obraz nie tyle odtwarza rzeczywistość, ale przedstawią ją przetworzoną, już z samego faktu zatrzymania sceny wizualnej w swoich ramach. I bynajmniej nie chodzi tutaj wyłącznie o treść przedstawienia, ale przede wszystkim o jego formę. W każdym razie ten sposób myślenia o obrazie zdominował niemal całą sztukę współczesną. W tym kontekście chciałbym też zwrócić uwagę na inne, niezwykle interesujące obrazy, które ilustrują różne rodzaje odstępstw od zasady dążenia do obsesyjnego potwierdzania treści za pomocą wszystkich trzech, interesujących nas kanałów analizy danych wzrokowych.
Kolorowanki według Picassa
Pierwszy przykład, to niemal drugi kraniec skali zgodności kształtów kodowanych przez kanał luminancji i kanały barw opozycyjnych. Przepiękny obraz Pabla Picassa, Matka i dziecko, przedstawia jego pierwszą żonę, Olgę Koklową z ich rocznym synem Paulem (ryc. 129).

Jest to znakomity przykład nonszalancji, z jaką artysta wypełnił barwami kontury narysowanych postaci. Kontur i barwne plamy przenikają się swobodnie, jakby nie były ze sobą niczym związane. Barwy zaledwie coś sugerują: zieleń – tło, błękit – bluzkę Paula, a nawet ich brak oznacza tyle, że sukienka Olgi jest po prostu jaśniejsza od żakietu. Jeszcze 30 lat wcześniej nikt nie zwróciłby uwagi na takie malowanie, uznano by je za szkic do obrazu, a nie skończone dzieło. A jednak ta niedbałość w położeniu barwnych plam bynajmniej nie przeszkadza, lecz przeciwnie, ożywia całą scenę.
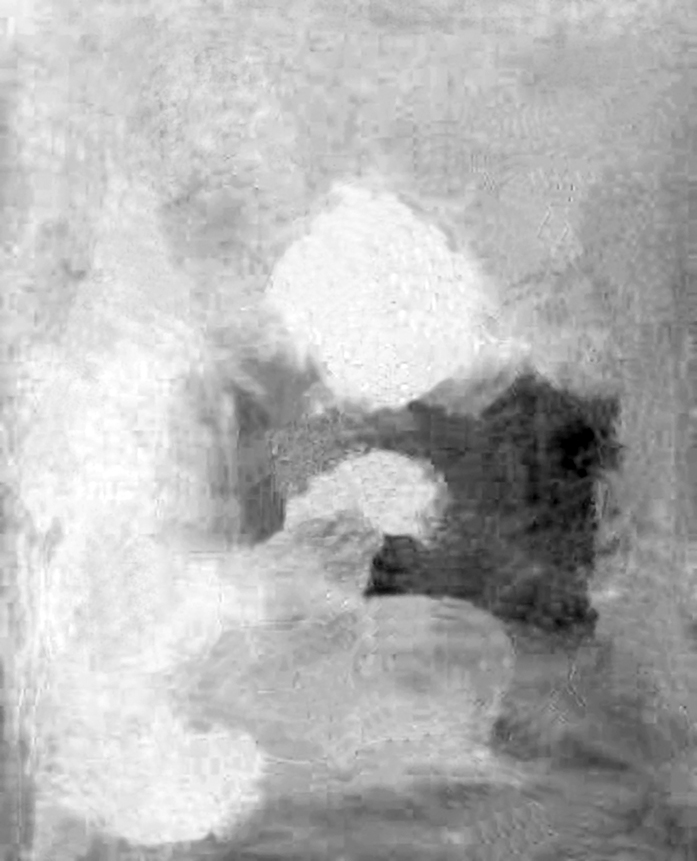
Po odseparowaniu rysunku od barwnych plam widać, jak dalece kontury matki i dziecka nie pokrywają się z krawędziami barwnych powierzchni (ryc. 130 A i B). Gdybyśmy wcześniej nie widzieli obrazu Picassa, to prawdopodobnie z trudem odgadlibyśmy, co przedstawiają te plamy. Jeszcze lepiej widać utratę kształtów kodowanych za pomocą barw po przefiltrowaniu ich przez kanały barw opozycyjnych (ryc. 130 C i D). Czarno-białe, niejednoznaczne plamy przypominają tablice projekcyjne, na które umysł obserwatora może rzutować niemal każde skojarzenie. To prawdopodobnie dzięki nim obraz Picassa nabiera nowych znaczeń i jest bardziej interesujący.

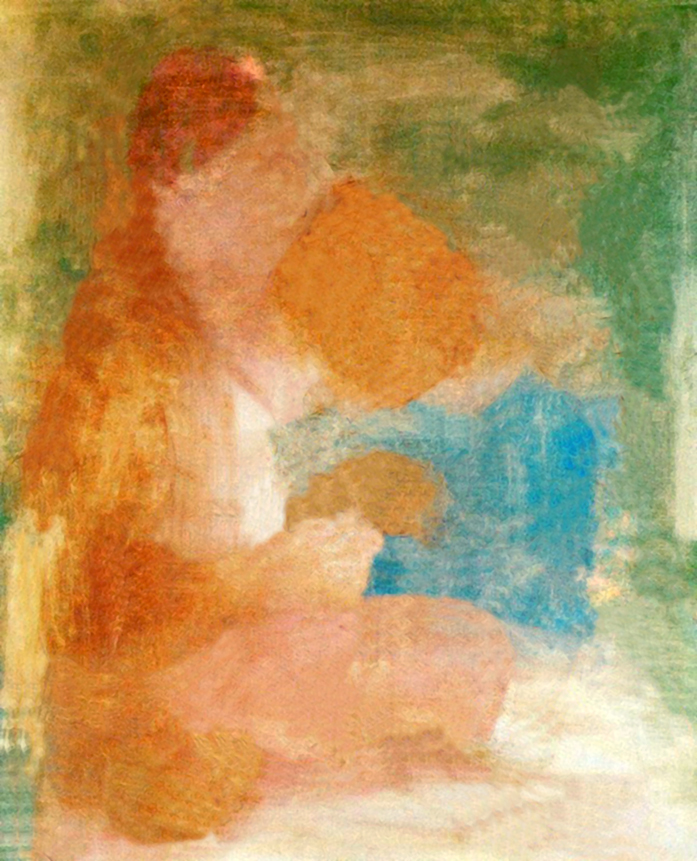

Henri i Amélie Matisse na życiowym wirażu
Kolejny przykład wybitnego dzieła malarskiego, na który chciałbym zwrócić uwagę to Kobieta w kapeluszu. Henri Matisse wystawił ten obraz w 1905 roku podczas Jesiennego Salonu w Paryżu (ryc. 131 A). Przedstawia on jego żonę, Amélię Parayre (ryc. 131 B) w wyrafinowanym kapeluszu.


Obraz Matisse’a podobnie, jak obraz Moneta, wywołał spore zamieszanie w ówczesnych kręgach artystów i znawców sztuki. Do tej pory nikt nie ośmielił się wystawiać obrazu namalowanego w taki sposób. To był policzek wymierzony akademikom i dobremu smakowi.
Ale co tak naprawdę rozdrażniło krytyków sztuki, że okrzyknęli Matisse’a i jego kolegów-malarzy bestiami (fauves) a Kobietę w kapeluszu „imponującym bohomazem”. „Pokazywany na paryskiej wystawie jest do dziś swoistym skandalem. Atakuje ostrymi kontrastami i niespokojną, rozbitą na kawałki powierzchnią. Kiedy pod przytłaczającym, ekstrawaganckim kapeluszem dostrzegamy w końcu twarz Amelii Matisse, uderza wrażenie niepewności i przygnębienia” pisze Anna Arno (2012) w artykule poświęconym Gertrudzie Stein i jej pasji gromadzenia dzieł Matisse’a. Ale czy tylko jest to twarz pełna „niepewności i przygnębienia”?
Rzut oka na twarz Amelii pod czerwonym kapeluszem potwierdza obserwacje Arno odnośnie do nastroju portretowanej modelki. Kiedy jednak zrównoważyłem jasność wszystkich barw na obrazie, a następnie odfiltrowałem pary barw opozycyjnych i zamieniłem wyniki tych filtracji na postać monochromatyczną, przed moimi oczyma pojawił się szokujący obraz. Oto spod ogromnego kapelusza wyłoniła się czaszka upiora, z pustymi oczodołami i fragmentami gnijącego ciała. Rozbicie obrazu na dwa kanały barw opozycyjnych ujawniły znacznie bardziej dramatyczną prawdę o wizerunku namalowanej kobiety niż wynikałoby to z jej powierzchownego oglądu (ryc. 132 A i B).


Czy ostre kolory, którymi Matisse pokrył twarz swojej żony były tylko ekstrawagancją malarza poszukującego nowych form wyrazu? A może raczej zostały one nie w pełni świadomie użyte przez niego w taki właśnie sposób, z powodu targających nim silnych, negatywnych emocji. Powodów nie brakowało, bowiem początek wieku był dla rodziny Maisse’ów prawdziwym koszmarem. W cztery lata po ślubie zostali całkowicie pozbawieni środków do życia wskutek bankructwa rodziców Amélie. Opuścili Paryż i zamieszkali na wsi. Krytyka całkowicie odrzucała wysiłki niemłodego już wówczas malarza. Matisse był bliski samobójstwa. Ostatnią deską ratunku okazało się rodzeństwo amerykańskich kolekcjonerów sztuki, Gertruda i Leo Stein. Dostrzegli oni wartość prac Matissa i zaczęli kupować jego obrazy.
Steinowie pomogli nie tylko Matissowi i jego rodzinie, ale także innemu wyjątkowemu artyście, który z ogromnym wysiłkiem przebijał się przez betonowy strop krytyki sztuki na początku XX wieku. Był nim Pablo Picasso.
Gdzie jest krzyż w scenie Ukrzyżowania?
Kończąc ten rozdział chciałbym przywołać jeszcze jeden przykład obrazu ilustrującego rozbieżności form kodowanych za pomocą opozycyjnych barw i luminancji. Chodzi o Ukrzyżowanie, obraz namalowany właśnie przez Pabla Picassa w 1930 roku (ryc. 133 A). Niemal wszystkie postaci i obiekty znajdujące się na nim, Picasso namalował ostrymi barwami. Tylko centralnie umieszczona scena tytułowego ukrzyżowania jest biała. Kontur krzyża oraz zniekształcone sylwetki Chrystusa i najprawdopodobniej Marii na białym tle prowokują do ich wypełnienia, jak w dziecięcej kolorowance albo witrażu.

Monochromatyczna wersja obrazu, podobnie, jak Kobieta w kapeluszu Henri Matisse’a, odzwierciedla położenie wszystkich obiektów, postaci i ich części w scenie, tak jak w obrazie barwnym (ryc. 133 B). Z tego punktu widzenia można uznać, że jaskrawe barwy są tylko ekstrawagancją malarza, kontestującego klasyczne standardy malarstwa przedstawieniowego. Dopiero obejrzenie tego dzieła przez pryzmat filtru czerwono-zielonego (ryc. 133 C) i żółto-niebieskiego (ryc. 133 D) ujawnia zaskakującą prawidłowość. W obu tych wersjach zanika centralna scena ukrzyżowania. Pozostaje rodzaj zasłony lub po prostu pustej powierzchni. Jest to sytuacja odwrotna w stosunku do efektu uzyskanego przez Claudie Moneta w Impresji. O ile tam, jasność słońca była bardzo podobna do jasności szaroniebieskiego tła i tylko barwy sygnalizowały jego obecność na obrazie, o tyle w dziele Picassa, kontrasty jasności pozwalają bez trudu zidentyfikować centralną scenę, ale już nie barwy.


